
La Société Française d’Ecologie (SFE) vous propose ce regard de Jean-François Ponge, Professeur émérite au Muséum National d’Histoire Naturelle, sur la dynamique et le fonctionnement des sols.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
Variations en sol majeur
par Jean-François Ponge,
Professeur émérite au Muséum National d’Histoire Naturelle
——-
Mots clés : sol, forêt, interactions, fonctionnement, écosystèmes, régénération forestière, vers de terre, formes d’humus, réseaux mycorhiziens, signaux chimiques
——–
Introduction
La Directive Cadre Européenne sur la Protection des Sols, qui devait établir les bases d’une protection durable des sols, a été officiellement retirée par la Commission européenne en Juin 2014, sous la pression des lobbies agricoles et industriels. Une initiative citoyenne européenne, People4Soil, portée par plus de 400 associations et en attente du recueil du million de signatures exigé par les règles communautaires (https://www.people4soil.eu/fr), invite la Commission européenne à présenter une proposition législative allant dans le sens de la prise en compte d’une des priorités majeures de notre millénaire. Mais pourquoi le sol est-il un enjeu aussi important et pourquoi certains craignent tant qu’on s’attache à sa protection ?

© Anne Teyssèdre
Longtemps utilisé comme un simple support des cultures, juste utile pour l’absorption de l’eau et des éléments minéraux, le rôle fondamental du sol dans le fonctionnement des écosystèmes terrestres – tels que champs et forêts – est aujourd’hui largement reconnu (cf. regards n°24, n°28, RE2). Mieux : avec ses réseaux d’interactions entre plantes, microbes et animaux, abritant l’essentiel de la biodiversité des milieux terrestres (Ponge 2003, 2013), le sol est considéré par certains auteurs comme un écosystème à part entière, emboîté dans l’écosystème forestier, prairial, etc. (De Ruiter et al. 1998, Ponge 2015). Au travers de trois exemples, nous allons voir que le sol est un élément-clé de l’équilibre dynamique des forêts tropicales humides et des forêts de montagne, et le lieu privilégié de l’intégration fonctionnelle des milieux terrestres.
Vertébrés frugivores et invertébrés du sol : une chaîne de processus au service de la dynamique des forêts tropicales
Les études menées en Guyane Française (Pouvelle et al. 2008, 2009, Dos Santos Neves et al. 2010, Feer et al. 2013) ont montré l’existence d’une chaîne de processus reliant les arbres, les vertébrés frugivores et les animaux du sol, assurant la pérennité des forêts tropicales humides. Longtemps connu comme un acteur essentiel de la régénération forestière, consommant les fruits charnus et transportant les graines loin des arbres parents et assurant ainsi leur dispersion à longue distance, le singe hurleur roux (Alouatta seniculus), le plus gros primate de la forêt guyanaise, est à l’origine d’une chaîne de processus impliquant le sol comme un acteur-clé de la régénération forestière.
Tous les soirs, après avoir fait une abondante provision de nourriture, les bandes de singes hurleurs se réunissent pour passer la nuit dans des arbres dont la hauteur dépasse souvent la canopée alentour (arbres dits « émergents »). Sous ces grands arbres dortoirs les singes défèquent et urinent. Ainsi le sol au pied des arbres émergents renferme une quantité extraordinaire de graines et s’enrichit en nutriments. Via des molécules odorantes, leurs déjections attirent de nombreuses espèces de bousiers, qui enfouissent en quelques minutes la matière fécale et les graines qu’elle renferme, à une faible profondeur, là où elles seront hors d’atteinte des nombreux prédateurs de surface tout en étant capables de germer lorsqu’une arrivée de lumière déclenchera leur germination (photoblastisme positif).
Une banque de graines* est ainsi constituée (une connaissance nouvelle pour les forêts tropicales), notamment pour les plus petites graines, à faibles réserves, qui vont trouver lors de leur germination une richesse en nutriments leur permettant une croissance rapide.
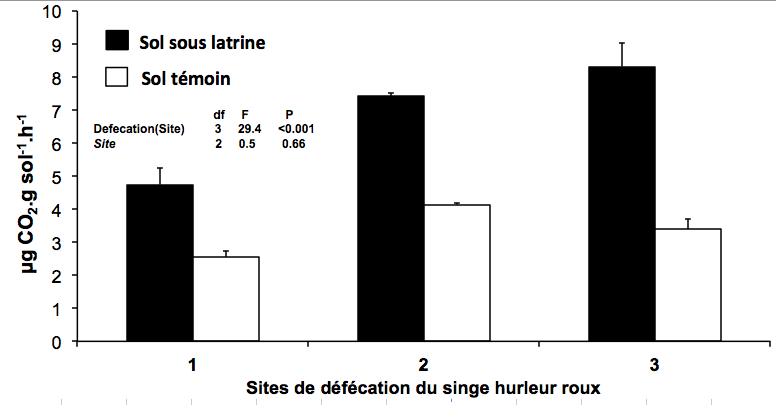
Figure 1. Mesure du métabolisme du sol dans le sol situé sous les latrines du singe hurleur roux et dans le sol témoin (distant des latrines).
© Jean-François Ponge & Nadia Dos Santos Neves, MNHN-CNRS.
La matière fécale des primates frugivores représente également une source de nourriture abondante et variée pour les vers de terre et les termites, organismes-ingénieurs des écosystèmes*, ainsi que pour la microflore et le système racinaire des arbres, stimulant ainsi le métabolisme du sol (cf. Figure 1). Il ne manque plus que la lumière pour assurer la levée de cette banque de graines, dont la survie ne peut cependant être assurée sur le long terme (contrairement aux forêts tempérées) en raison du climat chaud et humide.
Deux phénomènes vont contribuer à faire de ces dortoirs (également appelés latrines) des lieux privilégiés de la régénération de la forêt tropicale humide. D’une part, la banque de graines est alimentée en permanence, car les animaux utilisent régulièrement les mêmes arbres, d’autre part ces arbres de très grande taille ont plus de chances de tomber sous le poids des ans (sénescence, maladies) et/ou des intempéries. Leur chute éventuelle cause une trouée ou chablis* dans la forêt, où la lumière (verticale sous les tropiques) va permettre la germination et la levée rapide d’espèces végétales pionnières à croissance rapide. Ces espèces pionnières sont celles dont les graines se trouvent précisément là où le processus de reconstitution de la canopée peut se mettre en place rapidement et refermer les trouées en quelques années. Cette fermeture du couvert forestier permet la germination et la levée des espèces d’ombre, à grosses graines, germant très rapidement grâce à leurs réserves – espèces qui ne peuvent s’installer que sous le couvert des arbres. La boucle est ainsi fermée.

Canopée et arbres émergents de la forêt de Nouragues, Guyane Française
© Sandrine Pouvelle, MNHN-CNRS
Dynamique des populations de vers de terre et cycle des humus :
un élément-clé de la régénération des forêts de montagne
Les travaux effectués en Savoie (Bernier & Ponge 1994, Bernier 1996, 1997, Ponge et al., 1998) ont montré l’existence d’une répartition en mosaïque des formes d’humus* associée à la dynamique des communautés de vers de terre, en rapport étroit avec la régénération des forêts semi-naturelles d’épicéas (pessières savoyardes). Les sites de régénération sont de petites clairières (chablis) causées par la chute d’un ou plusieurs arbres âgés, ou bien les sites où les arbres ont été exploités selon le procédé traditionnel de la « cueillette ».
Dans ces sites, où le sol est couvert d’une végétation herbacée riche en nutriments, les communautés de vers de terre sont complètes – c’est-à-dire qu’elles comportent des espèces endogées, anéciques* et épigées – et participent à l’élaboration d’une forme d’humus appelée Mull*, à litière mince et bien aérée, propice à l’installation des jeunes semis de l’épicéa commun (Picea abies). Une fois cette installation terminée (le processus dure environ une vingtaine d’années à l’étage subalpin) le jeune peuplement va grandir, la communauté de vers de terre continuant à enrichir le sol en matière organique et en nutriments grâce à la présence d’une strate herbacée abondante et diversifiée.
Lorsque la canopée se referme, par coalescence des couronnes des arbres, le peuplement forestier devient un perchis* et la croissance rapide des arbres, jointe à l’appauvrissement de la litière du fait de la disparition de la strate herbacée, acidifie le sol et provoque un déséquilibre entre d’une part les besoins nutritionnels de la communauté animale, microbienne et végétale (arbres inclus), et d’autre part le recyclage des nutriments via la décomposition de la litière. Les populations de vers de terre s’appauvrissent en groupes fonctionnels* et les réseaux trophiques du sol passent d’un modèle fonctionnel basé sur les bactéries (lié au rôle d’ingénieur du sol des vers de terre) à un modèle basé sur les champignons, qui assure, essentiellement via l’activité des champignons mycorhiziens*, un circuit fermé des nutriments passant directement de la litière vers l’arbre. L’humus passe du Mull au Moder*, avec une faune dominée par les enchytréides* et les microarthropodes.
Contrairement aux anciennes idées, selon lesquelles cette évolution des formes d’humus était inéluctable sous les épicéas, on a pu montrer que cela ne constituait que la première partie d’un cycle où l’on voit, à condition que des sources de colonisation existent à proximité, et qu’on autorise les arbres à croître jusqu’à leur maturité, se reconstituer très progressivement des communautés fonctionnellement complètes de vers de terre (cf. figure 2 ci-dessous), avec un retour du Mull, avant même que les arbres parents meurent ou deviennent sénescents. Dans ces conditions, et seulement dans ces conditions, lorsque survient le chablis, l’humus est prêt à accueillir une nouvelle génération d’épicéas, le processus pouvant alors se poursuivre à l’infini.
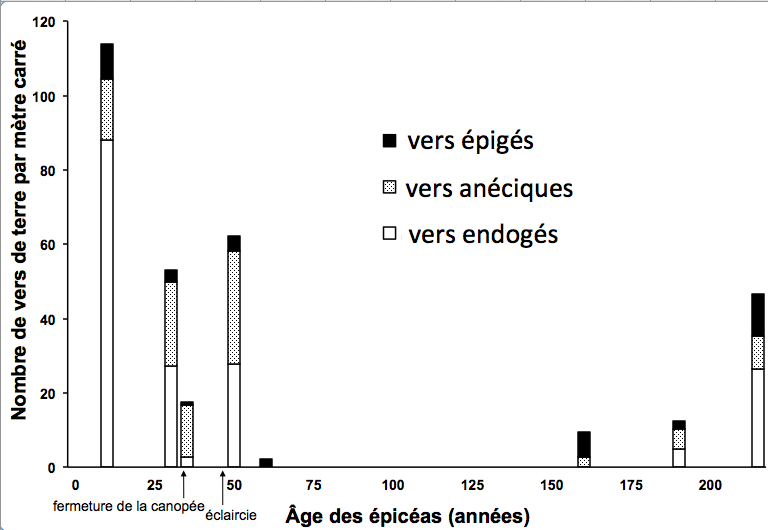
Figure 2. Abondance de trois groupes fonctionnels de vers de terre dans des peuplements d’épicéas d’âge croissant (forêt de Macot, Savoie, altitude 1500 m).
© Nicolas Bernier et Jean-François Ponge, MNHN-CNRS
Le sol des forêts de montagne et des forêts tropicales : une leçon pour la gestion durable de nos forêts
Dans les deux exemples cités, on a pu voir que les cycles observés faisaient appel à des acteurs indispensables, dont la disparition est susceptible de rompre les chaînes de processus observées. Les populations de singes hurleurs sont fortement affectées par la chasse à la « viande de brousse », partout où la pénétration de la forêt est facilitée par le développement du réseau routier et la croissance de l’urbanisation. La chasse constitue donc une menace majeure pour l’ensemble du processus, biodiversité des sols incluse bien entendu, et l’ensemble des processus sous son contrôle.
Dans le cas des forêts de montagne, c’est le mode de gestion hérité des forêts de plaine qui présente un risque majeur. Ainsi, une grande partie de la forêt communale de Macot-la-Plagne (en Savoie) est aujourd’hui gérée selon les principes de la sylviculture « nancéenne », avec des unités de gestion de plusieurs hectares d’un seul tenant, sans commune mesure avec le mode de fonctionnement par petites surfaces, lié à la dynamique des trouées, exposé ci-dessus. Or, pour que les vers de terre puissent circuler ‘horizontalement’ et reconstituer leurs populations lorsque les conditions locales le permettent, il faut des sources de recolonisation à proximité, donc des clairières ou du moins de très jeunes peuplements. Seule une gestion respectueuse de la mosaïque forestière naturelle des forêts de montagne, une gestion spécifique de ces milieux, le permet, ou plutôt le permettrait.
Les réseaux de champignons mycorhiziens, autoroutes de l’information dans les sols
Les associations de champignons et de racines (les mycorhizes*) sont connues depuis l’Antiquité et on a été longtemps fortement intrigué par l’absence de spécificité dans les associations observées, qu’il s’agisse des ectomycorhizes* (essentiellement à base de champignons basidiomycètes et quelques espèces d’ascomycètes) et des endomycorhizes* (essentiellement à base de champignons zygomycètes), les premières étant surtout présentes dans les sols forestiers à humus de type Moder, les secondes dans les sols forestiers à Mull et les formations herbacées (pelouses, prairies). Le réseau formé par les mycéliums des champignons mycorhiziens représente un réseau d’une longueur extraordinaire reliant entre eux les végétaux et les débris organiques, à la décomposition desquels certaines espèces de champignons mycorhiziens participent (Ponge 1990).
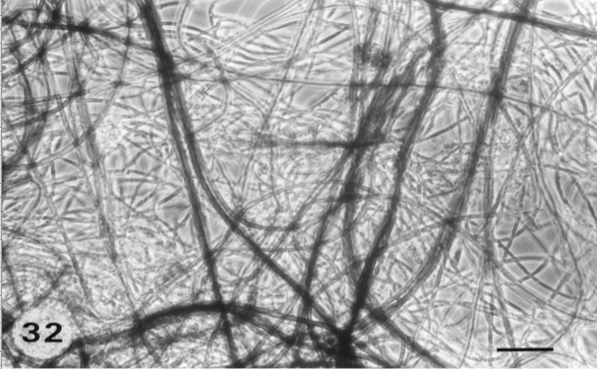
Mycélium extra-mycorhizien en réseau (deux espèces en mélange, l’une sombre l’autre claire), présent en abondance dans les couches de litière d’un peuplement de pin sylvestre. La connexion avec les racines de pin transformées (mycorhizes) a été vérifiée par l’observation microscopique.
© Jean-François Ponge, MNHN-CNRS
S’il est connu depuis longtemps que les mycéliums* des champignons mycorhiziens transportent des molécules organiques et des éléments minéraux sur d’assez longues distances, permettant des échanges entre plantes de la même espèce (y compris entre arbres « parents » et « enfants » en milieu forestier), seuls des travaux récents ont mis le doigt sur leur rôle dans les interactions fonctionnelles entre végétaux de différentes espèces, et donc sur leur fonction intégratrice au sein des communautés végétales (Selosse et al. 2006). Ces connaissances nouvelles sur le fonctionnement biologique du sol révolutionnent notre compréhension des lois qui régissent les assemblages d’espèces végétales et microbiennes (Bever et al. 2010). Dans le même ordre d’idées, les travaux portant sur les signaux échangés au sein du sol entre organismes animaux, végétaux et microbiens (Blouin et al. 2005) contribuent à modifier notre vision du sol et du rôle qu’il a pu jouer comme unité de sélection au cours de l’évolution (Rosenberg & Zilber-Rosenberg 2016), un domaine encore largement spéculatif mais largement porteur d’avenir.
Le sol a-t-il un avenir ?
Les trois exemples ci-dessus nous montrent que le sol joue un rôle fondamental dans l’intégration et la dynamique des milieux terrestres, qu’ils soient herbacés (prairies, champs) ou forestiers, et ce en l’absence de toute intervention humaine, depuis des millions d’années. L’homme en a tiré profit depuis des millénaires, contribuant à sa dégradation dès lors que des pratiques durables, issues souvent d’un savoir agronomique ancestral, ont été abandonnées. L’humanité va-t-elle être capable de préserver cette richesse ? On peut raisonnablement l’espérer, malgré les attaques incessantes des professionnels de l’agriculture intensive, pour qui le sol n’offre malheureusement aucune valeur ajoutée, donc ne génère pas de profit. Le sol, inscrit au Patrimoine Mondial de l’UNESCO ? Et pourquoi ne pas en rêver ?
—–
Glossaire
Anécique : Catégorie écologique de vers de terre effectuant des mouvements entre la surface, où ils se nourrissent de litière (notamment pendant le nuit), et la profondeur où ils excrètent la matière ingérée et la mélangent au sol minéral.
Banque de graines : Ensemble des graines enfouies dans le sol et susceptibles de germer lorsque les conditions deviennent favorables.
Chablis : Trouée formée par la chute naturelle d’un ou plusieurs arbres, dans une forêt.
Champignon mycorhizien : Champignon associé aux racines de certaines plantes, formant des associations étroites appelées mycorhizes.
Ectomycorhizes : Mycorhize où le champignon forme un manchon (appelé manteau) autour de la racine et ne pénètre que les interstices entre les cellules de la racine.
Endomycorhizes : Mycorhize où le champignon pénètre les cellules de la racine, sans former de manteau autour de celle-ci.
Enchytréides : Petits vers transparents, voisins des vers de terre, qui consomment comme eux de la matière organique mais dont la très petite taille ne permet pas son enfouissement en profondeur.
Forme d’humus : Type d’assemblage et de répartition de la matière organique dans un profil de sol.
Groupe fonctionnel : Groupe d’espèces assurant une même fonction au sein d’un écosystème.
Moder : Forme d’humus où la matière organique n’est pas directement incorporée à la matière minérale sous la forme d’agrégats organominéraux mais forme des horizons superposés où la litière est progressivement transformée.
Mull : Forme d’humus où la matière organique provenant de la litière est incorporée à la matière minérale sous la forme d’agrégats provenant de l’action d’organismes fouisseurs.
Mycélium : Ensemble de filaments de champignons formant un voile plus ou moins lâche, dans le sol ou à la surface des feuilles mortes.
Mycorhize : Racine modifiée par la symbiose avec un champignon (appelé champignon mycorhizien).
Organismes-ingénieurs des écosystèmes : Organismes dont l’activité modifie le milieu physique et les conditions de vie de nombreux autres organismes.
Perchis : Peuplement forestier dense formé d’arbres d’âge similaire et dans leur phase de croissance intense.
—–
Bibliographie
Bernier, N., 1996. Altitudinal changes in humus form dynamics in a spruce forest at the montane level. Plant and Soil 178, 1-28. https://www.researchgate.net/publication/225935564
Bernier, N., 1997. Fonctionnement biologique des humus et dynamique des pessières alpines: le cas de la forêt de Macot-la-Plagne (Savoie). Écologie 28, 23-44. https://www.researchgate.net/publication/233758152
Bernier, N., Ponge, J.F., 1994. Humus form dynamics during the sylvogenetic cycle in a mountain spruce forest. Soil Biology and Biochemistry 26, 183-220. https://www.researchgate.net/publication/46312511
Bever, J.D., Dickie, I.A., Facelli, E., Facelli, J.M., Klironomos, J., Moora, M., Rillig, M.C., Stock, W.D., Tibbett, M., Zobel, M., 2010. Rooting theories of plant community ecology in microbial interactions. Trends in Ecology and Evolution 25, 468-478. https://www.researchgate.net/publication/44682719
Blouin, M., Zuily-Fodil, Y., Pham-Thi, A.T., Laffray, D., Reversat, G., Pando, A., Tondoh, J., Lavelle, P., 2005. Belowground organism activities affect plant aboveground phenotype, inducing plant tolerance to parasites. Ecology Letters 8, 202-208.
De Ruiter, P.C., Neutel, A.M., Moore, J.C., 1998. Biodiversity in soil ecosystems: the role of energy flow and community stability. Applied Soil Ecology 10, 217-228. https://www.researchgate.net/publication/223924241
Dos Santos Neves, N., Feer, F., Salmon, S., Chateil, C., Ponge, J.F., 2010. The impact of red howler monkey latrines on the distribution of main nutrients and on topsoil profiles in a tropical rain forest. Austral Ecology 35, 549-559. https://www.researchgate.net/publication/232659529
Feer, F., Ponge, J.F., Jouard, S., Gomez, D., 2013. Monkey and dung beetle activities influence soil seed bank structure. Ecological Research 28, 93-102. https://www.researchgate.net/publication/242653997
Ponge, J.F., 1990. Ecological study of a forest humus by observing a small volume. I. Penetration of pine litter by mycorrhizal fungi. European Journal of Forest Pathology 20, 290-303. https://www.researchgate.net/publication/47629427
Ponge, J.F., 2003. Humus forms in terrestrial ecosystems: a framework to biodiversity. Soil Biology and Biochemistry 35, 935-945. https://www.researchgate.net/publication/222567430
Ponge, J.F., 2013. Plant-soil feedbacks mediated by humus forms: a review. Soil Biology and Biochemistry 57, 1048-1060. https://www.researchgate.net/publication/247161342
Ponge, J.F., 2015. The soil as an ecosystem. Biology and Fertility of Soils 51, 645-648. https://www.researchgate.net/publication/276090499
Ponge, J.F., André, J., Zackrisson, O., Bernier, N., Nilsson, M.C., Gallet, C., 1998. The forest regeneration puzzle: biological mechanisms in humus layers and forest vegetation dynamics. BioScience 48, 523-530. https://www.researchgate.net/publication/247844333
Pouvelle, S., Feer, F., Ponge, J.F., 2008. Topsoil as affected by dung deposition under resting places of red howler monkey (Alouatta seniculus). Pedosphere 18, 691-698. https://www.researchgate.net/publication/44709599
Pouvelle, S., Jouard, S., Feer, F., Tully, T., Ponge, J.F., 2009. The latrine effect: impact of howler monkeys on the distribution of small seeds in a tropical rain-forest soil. Journal of Tropical Ecology 25, 239-248. https://www.researchgate.net/publication/44567083
Rosenberg, E., Zilber-Rosenberg, I., 2016. Microbes drive evolution of animals and plants: the Hologenome Concept. mBio 7(2):e01395-15. https://www.researchgate.net/publication/299541433
Selosse, M.A., Richard, F., He, X.H., Simard, S.W., 2006. Mycorrhizal networks: des liaisons dangereuses? Trends in Ecology and Evolution 21, 621-628. https://www.researchgate.net/publication/6941351
Et ces regards et débats sur la biodiversité en ligne sur cette plateforme SFE :
Barot S et F. Dubs, 2012. Les écosystèmes du sol. Regards et débats sur la biodiversité, SFE, Regard n°28, 17 février 2012.
Couvet D. et A. Teyssèdre, 2016. Quelles politiques agricoles pour le 21e siècle ? Regards et débats sur la biodiversité, SFE, Regard n°68, 21 novembre 2016.
Doré T., 2011. La biodiversité, atout pour l’agriculture. Regards et débats sur la biodiversité, SFE, Regard n°24, 22 novembre 2011.
Dutoit T, 2013. L’ingénierie écologique. Regards et débats sur la biodiversité, SFE, Regard n°44, 5 avril 2013.
Etudiants de l’Institut Polytechnique La Salle – Beauvais, 2016. 1000 Kilogrammes sous vos pieds… Regards et débats sur la biodiversité, SFE, Regard étudiant RE2, 5 mai 2016.
Papy F. et I. Goldringer, 2011. La biodiversité des champs. Regards et débats sur la biodiversité, SFE, Regard n°21, 22 septembre 2011.
Thompson J. et O. Ronce, 2010. Fragmentation des habitats et dynamique de la biodiversité. Regards et débats sur la biodiversité, SFE, Regard n°6, 18 novembre 2010.
—
Article édité et mis en ligne par Anne Teyssèdre.
Merci pour cet article soulignant bien le rôle du sol dans l’écosystème. Les recherches pourraient être utilement complétées par les évolutions récentes en matière de modélisation mathématique, telles le modèle MOMOS qui permet de prédire l’écologie fonctionnelle des microorganismes et les échanges de carbone et d’azote (C et N) entre divers organismes vivants.
Merci pour ce rappel. Le modèle MOMOS est présenté ici:
http://horizon.documentation.ird.fr/exl-doc/pleins_textes/pleins_textes_7/b_fdi_59-60/010025752.pdf
Merci Jean-François pour ce bel article, précis et ouvert, comme d’habitude. Ta persévérance et la somme de tes observations patientes et perspicaces ont contribué à faire de notre communauté un pôle important et toujours à la pointe. Et c’est réconfortant de voir que la relève est active et forte!!
Très amicalement. Patrick
Merci pour ce bel article. Un peu dommage que la chute laisse l’impression d’une rêverie et n’ouvre pas sur un acte performatif. Un regard aboutissant à une pétition pour classer les sols au patrimoine mondiale de l’UNESCO ou vers un groupe de travail suivant cet objectif serait encore meilleur.
Pour moi qui crois que la culture ou la civilisation ne sont pas l’invention de l’homme mais celle naturelle du vivant, cette inscription me semblerait extrêmement sensée. Mais je pense que pour réaliser cette inscription de la meilleure façon, il faudrait d’abord, après la révision de 2004 des critères de l’UNESCO unifiant naturel et culturel (http://whc.unesco.org/fr/criteres/), maintenant mêler les critères immatériel et matériel. Ainsi les sols seraient protégés pour ce qu’ils sont : une pilier de notre civilisation, autant culturel que naturel, autant par l’esprit que par le territoire.
Oui Michel, mais je viens d’être informé par un collègue de Lille que le classement de l’ensemble des terrils du Nord pas de Calais au titre du Patrimoine de l’UNESCO (http://www.bassinminier-patrimoinemondial.org/) n’avait qu’une valeur symbolique et ne constituait en aucune sorte une protection. Ce terrils, aujourd’hui porteurs d’une richesse floristique et faunistique inégalée dans la région concernée, continuent à être exploités par des sociétés privées avec la bénédiction des édiles locaux.
Donc, non, l’UNESCO n’est pas la bonne solution (je ne le savais pas au moment de la rédaction de mon « regard »), seule la loi peut offrir une garantie de protection des sols…