
La Société Française d’Ecologie (SFE) vous souhaite une bonne année 2017 avec ce regard de Benoît FONTAINE, Benjamin BERGEROT, Isabelle LE VIOL et Romain JULLIARD, chercheurs au Centre d’Ecologie et Sciences de la Conservation (CESCO, MNHN), sur les impacts de l’urbanisation et du jardinage sur les communautés de papillons.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
Impact de l’urbanisation
et des pratiques de jardinage
sur les communautés de papillons
par Benoît Fontaine, Benjamin Bergerot, Isabelle Le Viol
et Romain Julliard,
chercheurs au CESCO, UMR 7204, MNHN
——-
Mots clés : Suivi de biodiversité, urbanisation, impacts, habitats, paysages, jardins, pratiques, papillons, communautés, écologie, méthodes, science participative
——–
Introduction
Les interactions multi-échelles sont fréquemment étudiées dans les recherches en écologie, car elles permettent de comprendre l’impact des processus à petite échelle sur les grands patterns, et inversement comment les facteurs qui jouent à grande échelle influent sur les dynamiques locales de la biodiversité (voir Peters et al., 2007). Les effets de ces interactions sont complexes, et l’importance relative des facteurs locaux et paysagers varie avec les espèces (taxons), selon par exemple leur degré de spécialisation à l’habitat ou leurs capacités de dispersion.
Dans ce contexte, les papillons de jour constituent un modèle biologique intéressant, non seulement du fait de leurs rôles dans les écosystèmes, mais aussi parce que leur cycle de vie court et leurs capacités de dispersion variées permettent de mesurer relativement facilement l’impact des variables environnementales. Ces caractéristiques sont bien entendu partagées par beaucoup d’autres groupes d’invertébrés, mais les papillons sont sans contexte le groupe d’insectes le mieux connu.

© A. Teyssèdre
Parmi les facteurs qui jouent un rôle dans la modification des patrons de biodiversité, l’urbanisation tient une place prépondérante (par ex Garaffa et al., 2009 ; Muratet et al., 2008). Les jardins, représentant des oasis vertes dans une matrice peu hospitalière, sont des ressources potentiellement importantes en ville, notamment car ils fournissent des ressources alimentaires pour les adultes dits imagos (Toms et al., 2010). L’étude présentée ici s’inscrit dans ce cadre, et s’intéresse aux impacts croisés de l’urbanisation et des caractéristiques des jardins sur les communautés de papillons communs en France. En s’appuyant sur un jeu de données à long terme collecté grâce à un programme de suivi participatif national, nous avons cherché à identifier les pratiques susceptibles de limiter l’effet délétère de l’urbanisation sur les communautés de papillons.
Données et protocole de recherche participative
Le jeu de données utilisé provient de l’Observatoire des Papillons des Jardins – OPJ – http://vigienature.mnhn.fr/page/operation-papillons ), programme de suivi ouvert à tous. Les participants à l’OPJ sont invités à identifier et compter les papillons dans leur jardin, à partir d’une liste fermée de 28 espèces et groupes d’espèces. Pour chaque espèce/groupe d’espèces, les relevés mensuels représentent le nombre maximum d’individus vus simultanément au cours du mois écoulé (mars à octobre). Par ailleurs, les participants remplissent un questionnaire décrivant les caractéristiques du jardin et de ses alentours (surface du jardin, présence d’éléments tels que pelouse, mare, friche ou potager, utilisation de pesticides, plantes présentes). Les données utilisées couvrent les années 2006 à 2012, pour 10 619 jardins.

Citron et abeille sur pois de senteur
© A. Teyssèdre
Complétant ces données locales par des données paysagères (estimées à l’aide de Corine Land Cover 2000), nous avons exploré l’impact de diverses variables environnementales -locales ou paysagères- sur trois variables descriptives des communautés de papillons de jardins : i) leur richesse spécifique mensuelle moyenne, c.-à-d. le nombre moyen d’espèces composant chaque mois ces communautés, ii) leur abondance globale mensuelle moyenne, c.-à-d. le nombre mensuel moyen d’individus de toutes espèces composant ces communautés iii) l’abondance mensuelle moyenne (c.-à-d. le nombre moyen d’individus par mois) de chaque espèce (ou groupe d’espèces) observée.
A l’échelle des paysages nous avons estimé à l’aide de Corine Land Cover la proportion de zone urbanisée (dite niveau d’urbanisation) et celle d’habitats naturels ou semi-naturels (i.e. hors zones urbanisées et agricoles) dans la commune de chaque jardin considéré.
Quant aux variables descriptives de l’habitat à l’échelle locale, fournies par les observateurs, il s’agissait de :
- La surface du jardin ;
- l’indice de naturalité du jardin : dans la description, les items suivants sont notés 1 si présents, 0 si absents : friche ; orties ; lierre ; ronces. L’indice de naturalité est la somme de ces scores ;
- l’indice d’offre en nectar du jardin : dans la description fournie, la présence de Buddleia, de centaurées, de lavande ou de ronces reçoit un score de 3; celles de valérianes, de trèfle ou de plantes aromatiques reçoit un score de 2, celle de géraniums de 1 (valeurs extraites de Bergerot et al. 2010). Pour chaque jardin, l’indice d’offre en nectar est la somme de ces scores;
- l’utilisation de pesticides (oui/non).
Quels impacts sur les communautés de papillons communs ?
Entre 2100 et 4000 jardins ont été suivis chaque année par les observateurs volontaires, ce qui représente plus de 100 000 relevés mensuels et environ 1,3 millions de papillons comptés.
Les analyses montrent que l’abondance globale et la richesse spécifique (ou diversité) moyennes des communautés de papillons, ainsi que l’abondance moyenne de 15 des 28 espèces/groupes d’espèces, varient inversement avec le niveau d’urbanisation. (En revanche, la proportion d’habitats naturels ou semi-naturels dans la commune n’a pas d’impact sur l’abondance ou la diversité totales moyennes.)
Parmi les variables locales, l’indice d’offre en nectar a un impact positif sur l’abondance totale des papillons, tandis que l’utilisation de pesticides, de façon attendue, a un impact négatif sur la diversité et l’abondance (cf. figure ci-dessous).
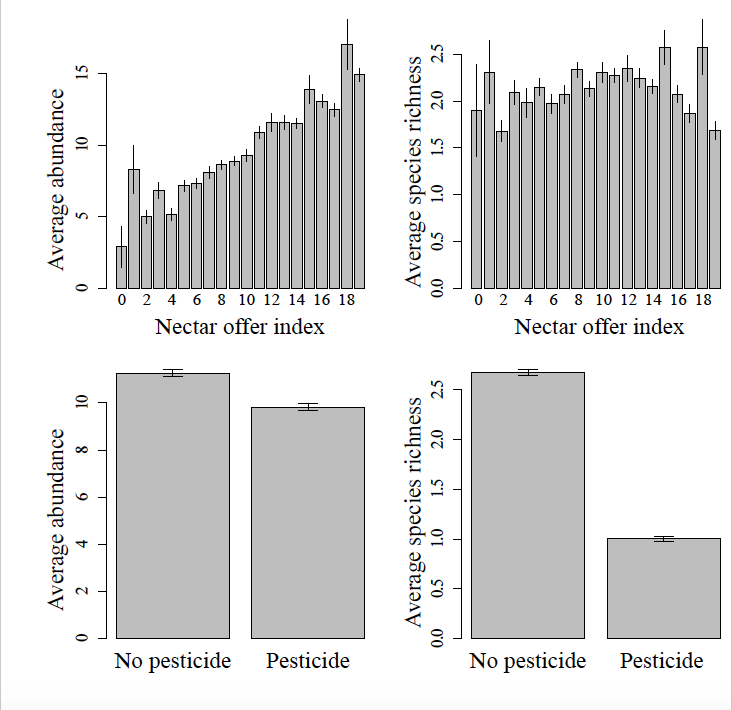
Fig.1 : Effets de l’offre en nectar et de l’utilisation de pesticides sur l’abondance et la diversité des communautés de papillons dans les jardins.
Au niveau spécifique, la plupart des espèces, y compris celles ayant des exigences strictes en matière de plante-hôte (sauf Argynnis paphia) sont impactées positivement par l’indice d’offre en nectar. Nous n’avons pas détecté d’impact de l’indice de naturalité sur l’abondance ou la diversité totale, bien qu’il y ait un effet sur onze espèces/groupes d’espèces. Enfin, la présence de certaines plantes, notamment les orties et les crucifères, influe positivement sur l’abondance des papillons, notamment ceux pour lesquels il s’agit des plantes-hôtes des chenilles.
Plusieurs interactions entre variables locales et paysagères ont un impact sur les papillons : l’effet positif de la surface du jardin sur l’abondance des papillons est plus important dans les zones très urbanisées, où ces jardins sont les seuls habitats favorables pour ces insectes. Ce résultat est confirmé par l’effet positif de la surface du jardin moins important lorsque le paysage environnant présente de grandes zones d’habitats naturels ou semi-naturels, qui sont également favorables aux papillons. Mais le résultat le plus remarquable réside dans le fait que pour l’ensemble des espèces/groupes d’espèces, la réponse au niveau d’urbanisation et celles à l’indice de naturalité et à l’indice d’offre en nectar sont corrélées négativement, ce qui signifie que les espèces réagissant le plus négativement à l’urbanisation sont celles qui bénéficient le plus de bonnes pratiques de jardinage – jardins plus naturels et/ou offrant davantage de ressources alimentaires pour les imagos (cf. Fig.2 ci-dessous).
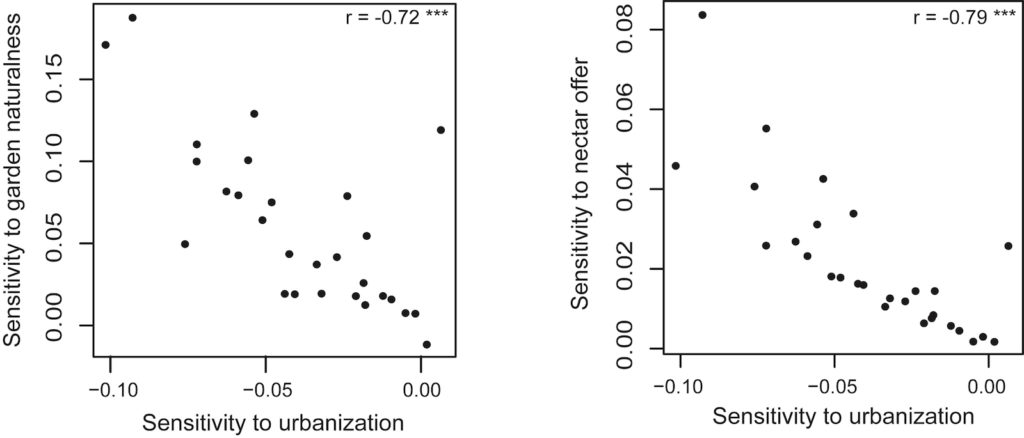
Fig.2 : Liens entre réponse à l’urbanisation et réponse à l’offre en nectar dans les jardins. Chaque point représente une des espèces/groupes d’espèces suivis. Les espèces les plus sensibles à l’urbanisation (partie gauche du graphe) sont celles qui bénéficient le plus des pratiques de jardinage (partie haute du graphe).
Les jardins en ville, refuges pour les papillons
Cette étude montre donc que l’impact négatif de l’urbanisation peut être limité localement par les caractéristiques du jardin et les pratiques de jardinage (types de plantes présentes ou utilisation de pesticides) : des pratiques prenant en compte les besoins des papillons sont efficaces même dans les zones très urbanisées, où les jardins représentent des refuges pour les espèces sauvages. Bien entendu, les communautés de papillons seront généralement moins abondantes et diversifiées en milieu urbain qu’en milieu rural. Les espèces spécialistes avec de faibles capacités de dispersion seront moins présentes en ville, quelles que soient les pratiques au niveau local. Mais les bonnes pratiques permettent de limiter l’impact négatif du paysage, un résultat important pour les gestionnaires de parcs et les jardiniers amateurs soucieux de l’environnement.

Morosphinx et valériane dans un jardin
© Eckard Berberich
Dans notre étude, le fort effet positif de l’offre en nectar sur l’abondance et la diversité des papillons, y compris pour les espèces spécialistes en termes de plante hôte, corrobore le fait que les jardins sont des sources de nourriture pour les papillons adultes, et il est probable que cette offre en nectar détermine la capacité de charge des jardins pour les communautés de papillons. Le fait que l’indice de naturalité du jardin n’ait pas d’effet sur l’abondance globale et le nombre moyen d’espèces (richesse spécifique) va dans le même sens. Cela étant, comme certaines espèces réagissent de façon significative à la naturalité du jardin, la composition de la communauté de papillons est influencée par ce descripteur, et les pratiques ont donc le potentiel de limiter l’homogénéisation biotique des milieux (cf. le Regard n°16 de Joanne Clavel sur cette plateforme SFE).
Il est probable qu’à niveau d’urbanisation égal, un indice de naturalité plus élevé bénéficiera à certaines espèces qui pourront se reproduire dans le jardin, où elles trouveront leurs plantes-hôtes (par exemple des Apiaceae pour le machaon Papilio machaon ; des Poaceae pour les demi-deuils Melanargia spp, les hespérides orangées, les Lasiommata, le myrtil Maniola jurtina, le procris Coenonympha pamphilus, le silène Brintesia circe et le tircis Pararge aegeria ; les chardons pour la belle-dame Vanessa cardui ; ou les orties pour la petite tortue Aglais urticae, le vulcain Vanessa atalanta, le paon du jour Inachis io et le robert-le-diable Polygonia c-album).

Jardin partagé, Paris © A. Teyssèdre
Epilogue: Les multiples atouts des observatoires citoyens
Ces résultats montrent également la puissance d’un programme de suivi de la biodiversité basé sur la collecte de données par de nombreux observateurs bénévoles, sans exigence d’expertise. Lorsqu’il s’agit de réaliser des analyses sur de gros jeux de données concernant la biodiversité, la science participative a de grands atouts (cf. Regard n°11). Ainsi, bien que représentant une grande proportion des espaces verts en ville, les jardins privés sont très peu étudiés à grande échelle du fait de la difficulté d’accès à propriétés privées en grand nombre : la science participative permet de contourner cet obstacle puisque ce sont les jardiniers eux-mêmes qui collectent les données.
D’autre part, les programmes de suivi impliquant des volontaires permettent de collecter des données à de grandes échelles spatiales et temporelles, avec une couverture qui serait impossible à atteindre en faisait appel à des professionnels, simplement par manque de main-d’œuvre disponible (et même si la main-d’œuvre existait, le coût serait prohibitif – Levrel et al., 2010). Enfin, la participation à de tels programmes implique une prise de conscience environnementale, et potentiellement un changement de comportement des observateurs vis-à-vis de la biodiversité (Couvet et al., 2008).
—–
Ce regard est une version vulgarisée, adaptée pour cette plateforme SFE, de l’article de B. Fontaine et al. ‘Impact of urbanization and gardening practices on common butterfly communities in France’, paru dans la revue Ecology and Evolution en octobre 2016.
—–
Site web de l’Observatoire des Papillons des Jardins (OPJ) : http://vigienature.mnhn.fr/page/operation-papillons
—–
Bibliographie
Bergerot B., Fontaine B., Renard M., Cadi A. & Julliard R., 2010. Preferences for exotic flowers do not promote urban life in butterflies, Landscape and Urban Planning 92(2): 98-107.
Couvet D., Jiguet F., Julliard R., Levrel H. & Teyssèdre A., 2008. Enhancing citizen contributions to biodiversity science and public policy, Interdisciplinary Science Reviews 33(1):95-103.
Fontaine B., Bergerot B., Le Viol I. & Julliard R., 2016. Impact of urbanization and gardening practices on common butterfly communities in France. Ecology and Evolution 6: 8174–8180.
Garaffa P. I., Filloy J. & Bellocq M. I., 2009. Bird community responses along urban-rural gradients: Does the size of the urbanized area matter?, Landscape and Urban Planning 90(1-2):33-41.
Levrel H., Fontaine B., Henry P. H., Jiguet F., Julliard R. & Kerbiriou, C., Couvet, D., 2010. Balancing state and volunteer investment in biodiversity monitoring for the implementation of CBD indicators: A French example, Ecological Economics 69 (7) : 1580-1586. <10.1016/j.ecolecon.2010.03.001>
Lizée M.-H., Tatoni T. & Deschamps-Cottin M., 2015, Nested patterns in urban butterfly species assemblages: respective roles of plot management, park layout and landscape features, Urban Ecosystems 19: 205. http://doi.org/10.1007/s11252-015-0501-5
Muratet A., Porcher E., Devictor V., Arnal G., Moret J., Wright S. & Machon N., 2008. Evaluation of floristic diversity in urban areas as a basis for habitat management, Applied Vegetation Science 11(4):451-460.
Pandit S. N., Kolasa J. & Cottenie, K., 2009. Contrast between habitat generalists and specialists: an empirical extension to the basic metacommunity framework, Ecology 90(8):2253-2262.
Peters D. P. C., Bestelmeyer B. T. & Turner M. G., 2007. Cross-scale intercations and changing pattern-process relationships: consequences for system dynamics, Ecosystems 10:790-796.
Toms M. P., Humphreys L., Kirkland P., 2010. Monitoring butterflies within an urbanised landscape: the role of garden butterfly populations in a wider context, in: The 2010 target and beyond for Lepidoptera, 26-28 March 2010 (M. Warren, J. Dover, eds.), Reading, UK, pp. 35.
Et ces regards et débats sur la biodiversité en ligne :
Clavel J., 2011. L’homogénéisation biotique. Regards et débats sur la biodiversité, SFE, Regard n°16, 18 avril 2011.
Clergeau P., 2010. Biodiversité urbaine : De l’inventaire naturaliste au fonctionnement écologique. Regards et débats sur la biodiversité, SFE, Regard n°8, 16 décembre 2010.
Julliard R., 2010. Regard sur une perruche. Regards et débats sur la biodiversité, SFE, Regard n°2, 25 septembre 2010.
Julliard R. et F. Jiguet, 2011. La biodiversité face au réchauffement climatique. Regards et débats sur la biodiversité, SFE, Regard n°22, 11 octobre 2011.
Teyssèdre A. et D. Couvet, 2011. Biodiversité et science participative, de la recherche à la gestion des écosystèmes. Regards et débats sur la biodiversité, SFE, Regard n°11, 6 février 2011.
Thompson J. et O. Ronce. Fragmentation des habitats et dynamique de la biodiversité. Regards et débats sur la biodiversité, SFE, Regard n°6, 18 novembre 2010.
Article édité et mis en ligne par Anne Teyssèdre.
——
Démarche intéressante que d’impliquer les citoyens dans un projet scientifique sur la conservation de la biodiversité en milieu urbain.
Deux questions à vous poser:
1-Avez-vous pu comparer ces résultats (impliquant les sciences participatives) avec ceux obtenus à partir de données produites par des professionnels selon une méthodologie protocolée mais forcément sur moins de sites?
2-Avez-vous engagé des actions de transfert des résultats de cette étude vers la société et si oui comment?
Merci pour vos réponses.
Bonjour,
Nous n’avons pas comparé ces résultats avec ceux obtenus à partir de données collectées par des professionnels, par manque de telles données. En revanche, nous avons testé la fiabilité des données protocolées obtenues par les citoyens en comparant les tendances à long terme ou la phénologie issues des données « citoyens » avec celles issues de données collectées par des naturalistes spécialistes des papillons. Ces comparaisons montrent que les données produites par les citoyens sont fiables, les résultats sont similaires à ceux obtenus par les lépidoptéristes. Cette fiabilité autorise à mettre en place des études comme celle présentée ici, études qui seraient très difficiles, voire impossibles à effectuer à cette échelle sans faire appel aux sciences participatives.
Nous communiquons sur les résultats obtenus à partir des observatoires participatifs par des communiqués de presse repris dans les médias, et directement auprès des participants à travers des bulletins d’information et au cours de séminaires auxquels ils participent. Et maintenant par le biais des Regards de la SFE !
Cordialement
Bonjour Benoît et al. 🙂
Votre article/regard me paraît très intéressant, notamment parce qu’il montre que l’abondance des communautés de papillons (en nombre total d’individus) et leur richesse spécifique (en nombre d’espèces) varient de façon très différente avec les variables explorées. Ainsi l’abondance des papillons est largement déterminée par l’offre en nectar des jardins (qui peut donc être considérée comme une proxy pour leur capacité de charge), mais peu sensible aux pesticides, tandis que c’est l’inverse pour la richesse spécifique (cf. la Fig.1).
Cependant, si l’on admet que l’abondance des communautés de papillons est déterminée par l’offre locale en nectar, le faible impact mesuré des pesticides sur l’abondance des communautés de papillons m’intrigue. (Ou en d’autres termes, la combinaison des deux schémas de gauche, figure 1, ne me semble pas facile à interpréter.) Est-ce que les pesticides n’ont qu’un très faible impact sur l’offre globale en nectar, via les plantes résistantes aux pesticides et riches en nectar? (Dans ce cas, avez-vous exploré l’impact sur les plantes des pesticides utilisés?) Ou bien s’agit-il de l’impact résiduel des pesticides sur l’abondance des communautés (de papillons), à offre en nectar constante?
Bonjour,
L’action des pesticides sur les communautés de papillons dans les jardins est complexe : comme nous l’avons montré par ailleurs (Muratet & Fontaine, 2015), si les insecticides et les herbicides utilisés dans les jardins ont un impact négatif sur les papillons (impact direct pour les insecticides, indirect pour les herbicides qui limitent les ressources des papillons), l’utilisation de fongicides, d’anti-limaces et de bouillie bordelaise est corrélée à une plus grande abondance de papillons. Nous faisons l’hypothèse que ce résultat inattendu pourrait être dû au fait que ces trois classes de pesticides protègent les plantes contre des organismes qui les affaiblissent habituellement, et favorisent une production de nectar plus importante, et donc une plus grande abondance d’insectes floricoles.
Dans le présent article, le but n’était pas d’étudier en particulier l’impact des pesticides sur les papillons : tous les types de pesticides ont donc été regroupés, et le design de l’étude ne permet pas ici de démêler les effets complexes et souvent indirects des pesticides utilisés par des jardiniers amateurs.
Bonjour Benoît,
et merci pour cette réponse, qui explique en effet le faible impact observé de l’ensemble des pesticides utilisés sur l’abondance des papillons.
Cependant, dans ce cas, le fort impact de ces mêmes pesticides sur la richesse des communautés de papillons (en nombre d’espèces) suggère que non seulement les herbicides, mais aussi les pesticides augmentant l’offre en nectar -tels que fongicides et ‘anti-limaces’-, réduisent la richesse de ces communautés.
Est-ce le cas? = Y a-t-il des résultats en ce sens? Et si oui, comment interpréter ces résultats?
NB: Si c’est le cas, une interprétation possible est que les pesticides non herbicides, tels que fongicides et anti-limaces, favorisent -en augmentant l’offre en nectar- un petit nombre d’espèces de papillons généralistes, aux dépens d’un plus grand nombre d’espèces spécialistes.
Y a-t-il d’autres résultats/observations en ce sens?
Bien cordialement,
Anne
Bonjour,
Il a été montré chez les oiseaux que l’augmentation des doses de pesticides pour l’agriculture est corrélée avec une augmentation de la proportion de généralistes (Chiron et al., 2014, Agriculture Ecosystems and Environment), un résultat cohérent avec l’effet connu de l’homogénéisation des communautés induite par l’intensification des pratiques. Il est effectivement probable que ce résultat s’applique aussi aux insectes.