La Société Française d’Ecologie et d’Evolution (SFE2) vous propose ce Regard de Patrick Lavelle, Professeur d’écologie émérite à l’IESS, sur l’écologie des sols et la recherche.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
L´Ecologie des sols face aux défis du siècle
Patrick Lavelle
Professeur Emérite, Institut d´Ecologie et des Sciences Environnementales (IEES-BIODIS),
Sorbonne Université, Paris. Membre Correspondant de l´Académie des Sciences
Regard RO20, édité par Anne Teyssèdre et Jérôme Cortet
—-
Mots-clés : Sols, fonctionnement, organismes ingénieurs, mutualisme, auto-organisation,
agriculture, gestion durable
—–
- Résumé
- Introduction
- Une écologie qui prend en compte la spécificité du sol et des interactions entre ses habitants
- De la facilitation à l’auto-organisation
- L’écologie du sol au service du futur : ingénierie biochimique et dynamique de l’auto-organisation
- Une communication scientifique plus détendue
- Du dualisme au monisme ?
- Le mot de la fin, avec Darwin
- Glossaire
- Annexe : Exercice proposé aux étudiants
- Remerciements
- Bibliographie
- Regards connexes
- Forum de discussion sur ce regard
——
« Les philosophes post-modernes suggèrent que la connaissance scientifique, en tant que construction sociale, est une construction subjective qui dépend de la langue et de la culture des scientifiques impliqués ». (Jagers op Akkerhuis, 2015)
Résumé
L´Ecologie du sol est populaire depuis quelques années, avec la publication d´ouvrages collectifs (Orgiazzi et al., 2016 ; FAO, 2020), un nombre important d´articles publiés dans les grandes revues, la décennie des sols et le programme Global Soil Biodiversity Initiative qui organise des évènements très suivis. Nous analysons les raisons pour lesquelles elle est restée aussi longtemps discrète, ne publiant que dans des revues spécialisées à faible impact, incapable d´influer le cours de la science du sol comme de celui de l´écologie générale. Nous soutenons l´idée que les écologistes du sol utilisant les paradigmes inadaptés de la « mainstream Ecology » ont en grande partie oublié les deux conditions qui auraient dû inspirer leur démarche théorique : 1. la vie dans les sols nécessite un aménagement constant et couteux en énergie du milieu pour créer les habitats et les conditions pour que l´eau et l´air y circulent et que s´y produisent les nombreux services écosystémiques des sols ; 2. que c´est le lieu du recyclage des 80% de la production primaire, qui une fois morte, échappe aux chaînes trophiques* classiques. L’exploitation par les organismes du sol de ce type de ressources nécessite des interactions différentes des classiques chaînes trophiques et de leur régulation par la compétition.
La représentation du sol par la théorie de l´auto-organisation élaborée au fil de trente années de recherches dans notre groupe met au premier rang les interactions positives entre organismes de tailles différentes – micro- et macro-organismes – et le rôle particulier des organismes ingénieurs* dans le fonctionnement du sol. Il est intéressant de se demander pourquoi cette approche a été celle d´une minorité de scientifiques, à dominante française, et pourquoi l’auto-organisation des sols n´est pas reconnue par tous comme la base théorique de la discipline.
Nous terminons ce regard en discutant dans quelle direction doit s´orienter la recherche et la méthodologie de diffusion des résultats et quels nouveaux paradigmes permettront au mieux d´accompagner le vaste mouvement de l´agriculture durable qui nécessite l´appui concret de notre discipline.
Introduction
L´écologie du sol a été longtemps limitée par la difficulté technique à explorer un milieu complexe, pauvrement renseigné par les taxonomistes et longtemps confinée a des revues sans grand impact et à court de paradigmes adaptés. L´ ignorance dans laquelle la tient la science du sol a été souvent signalée (Lavelle, 2000; Bottinelli et al., 2015 ; Filser et al., 2016). Notre discipline émerge enfin, avec la publication d´ouvrages multi auteurs importants (Orgiazzi et al., 2015 , https://www.researchgate.net/publication/303699867_Global_Soil_Biodiversity_Atlas#fullTextFileContent et FAO, 2020; http://www.fao.org/soils-portal/soil-biodiversity/en/) et l´existence d´un projet fédérateur, la Global Soil Biodiversity Initiative (www.globalsoilbiodiversity.org), tandis que la Décennie mondiale pour la restauration des écosystèmes décrétée par l´ONU en 2021 (https://www.iuss.org/index.php?article_id=588) devrait utiliser ces résultats (voir aussi les Regards 28, E2, 70, 89 et 99).
Le sol, enfin reconnu comme une ressource importante, à la base de 16 des 24 services écosystémiques reconnus par le Millennium Ecosystem Assessment (www.millenniumassessment.org), est en grand danger et les options de gestion proposées pour y remédier interpellent les scientifiques.
C´est le moment de nous demander si les bases théoriques sur lesquelles nous nous appuyons sont suffisamment consensuelles, claires et opérationnelles pour guider l´évolution attendue. Le moment aussi peut être, de les repenser et de les élargir pour être à la hauteur du défi présent des sociétés occidentales, fruit de leurs pressions croissantes sur l’environnement : changer radicalement leur projet d´humanité, hérité du siècle des lumières.
La vision cartésienne duale du monde, séparant l´esprit de la matière, a orienté la science vers une approche résolument matérialiste, inspirée par l´idéologie dominante du moment (Boucher et al., 1982 ; Descola, 2005). L´écologie avec une approche « mécaniste » a ainsi décrit avec succès le fonctionnement et la structure des écosystèmes, énoncé des lois et des paradigmes qui ont permis des avancées indéniables. Le modèle qui décrit le sol comme un système auto-organisé, où les organismes ingénieurs jouent un rôle prépondérant, est notre contribution à cet effort (Lavelle et al., 2016). Il a été élaboré au fil de la trentaine de thèses de doctorat que j´ai eu le privilège de co-encadrer.
Nous étions des écologues sensoriels (« les écologistes en sac à dos » comme nous appelait avec dédain André Mariotti, par ailleurs un grand scientifique qui nous a beaucoup fait progresser en nous ouvrant l´accès aux techniques isotopiques), rarement favorisés dans l´attribution des bourses de thèse. Les qualités d´observateur et l´aptitude à se mouvoir et à collecter des informations de qualité dans des milieux difficiles n´ont jamais vraiment été considérées comme importantes pour l´évaluation des étudiants et des chercheurs. Ma proposition d´inscrire la course d´orientation et le trail dans le bagage des étudiants en écologie n´a jamais reçu l´attention qu´elle mérite. Pourtant, la qualité des données collectées, à l´heure du big data (James et al., 2021), dépend aussi de l’aisance du chercheur à se mouvoir dans le milieu physique et social dans lequel il évolue et cet aspect qui se travaille, ne devrait pas être négligé !!
L’Institut de Recherche pour le Développement (IRD), institution ouverte sur le monde et les partenariats internationaux furent alors notre principal soutien. La priorité pour nous était l´observation des patrons du terrain dans de grands programmes d´échantillonnage et d´observations sur le terrain, suivant des protocoles stricts permettant l´usage d´outils statistiques avancés. L´expérimentation -dans les conditions les plus proches que possible de la nature- et la modélisation expliquaient les patrons observés sur le terrain. Combien de tonnes de sols, de centaines de sites de dizaines de pays différents, avons-nous soigneusement décortiqués à la recherche de la macrofaune, observant au passage le sol et ses agrégats, y lisant mille et une histoires de vie et analysant chaque processus à travers la vie et les interactions des organismes vivants (Figure 1) ?

Figure 1. Photo et légende : P. Lavelle
Une écologie qui prend en compte la spécificité du sol et des interactions entre ses habitants
L´essor de l´écologie du sol a sans doute été retardé par l´oubli général, dans les approches conceptuelles dominantes, des trois contraintes extrêmes imposées par le sol, à savoir la difficulté à se déplacer dans un environnement compact, l´alternance de phases aquatiques et aériennes dans l´espace poreux et, surtout, la difficile nature des ressources disponibles (Lavelle et Spain, 2001).
Le sol n´est pas un milieu où les prédateurs, les proies et les « ingénieurs de l’écosystème»* sont en contact permanent (Erktan et al., 2020) et se conforment aux prévisions des modèles mathématiques initiés par Lotka et Volterra il y a presque 100 ans (Volterra, 1931). Ce n´est pas non plus un milieu où la valeur adaptative des espèces se mesure forcément à leur compétitivité dans l´accaparement des ressources (Price, 1984). Ces ressources, qu´elles soient de « mauvaise » qualité (sensu Swift et al., 1979) comme la matière organique et la litière, ou de bonne qualité, mais distribuées de façon sporadique dans le temps et dans l´espace comme les exsudats racinaires, ne sont exploitables que grâce aux relations mutualistes entre organismes, d’une grande diversité dans le sol (Lavelle et Spain, 2001).
Le sol est justement le lieu du recyclage des 80% de la matière qui échappent aux réseaux trophiques* « classiques », épigés. Appliquer aux communautés endogées les dogmes consacrés du milieu aérien où chaînes trophiques et compétition organiseraient le partage harmonieux des ressources et entretiendraient la biodiversité, c´est oublier ces deux réalités incontournables :
- Sans l´activité biologique, les interactions et le mutualisme entre les organismes ingénieurs* (macro-invertébrés et racines) et les microorganismes et l´ énorme investissement énergétique des plantes qui injectent dans le sol 20% du produit de la photosynthèse sous forme d´exsudats, le sol n´est plus le milieu à la fois poreux et agrégé que nous observons, mais une brique compacte impropre à la vie.
- Les ressources disponibles dans le sol sont de la matière organique morte de faible valeur nutritive ou des ressources solubles, plusieurs tonnes par ha d’exsudats racinaires inoculés directement dans le sol, ressource de bonne qualité mais dispersée de façon imprévisible aux pointes des racines en croissance.
Oubliant le Darwin qui en 1881 s´extasiait, au-delà de toute considération scientifique matérialiste, de l´extraordinaire contribution des vers de terre á la création des sols (Gatineau, 2018) , les écologues ont été incités à suivre, et peut être même à surinterpréter, le Darwin des pinsons et les dogmes de l’ écologie « mainstream » (Barot et al., 2007; Thakur et al., 2020), influencés peut être par l´idéologie dominante de l´époque, comme le suggérait Kropotkin (1902) il y a plus d´un siècle. L´agacé « Ah ! Lavelle et ses vers de terre » combien de fois murmuré dans les débats contradictoires ici et là, auraient pu être avantageusement remplacé par un « Ah ! Darwin et ses vers de terre » plus approprié.
J´ai toujours combattu la vision –pour moi néolibérale et bouchère, pour le dire crument- voulant que les chaînes trophiques et la compétition déterminent le fonctionnement du sol (Nielsen et al., 2015), indépendamment de toute considération sur la physique du sol, sa dynamique et ses contraintes, de la difficulté à circuler dans ce milieu compact et de la faible qualité des ressources dont l’assimilation par digestion impose une coopération entre micro- et macro-organismes. Celle-ci se fait par un mode de digestion mutualiste entre les bactéries ingérées avec le sol et le ver qui crée pour elles des conditions très favorables à leur activité dans son intestin (Trigo et al.,1992).
Nous mesurions dans nos études que l´équivalent d’une couche de 10 cm de sol pouvait passer dans les intestins des vers tous les ans (Lavelle, 1978) et je demandais : que se passe-t-il quand ils ingèrent le réseau trophique, localisé dans les petits agrégats et les pores de taille intermédiaire que vers endogés et termites mangeurs d’humus (humivores) consomment sans modération ? Les travaux de Johnny Boyer montraient que les nématodes du sol ne ressortent pas indemnes d´un tel voyage, qu´ils soient directement digérés, qu´ils ressortent désorientés après qu´une bactérie ait détruit leurs chémorécepteurs… ou que l´éclosion des œufs soit accélérée, faisant naitre des nématodes dans la structure hostile et compacte d´un turricule d´où ils ne pourraient pas ressortir (Boyer et al., 2013).
De la facilitation à l’auto-organisation
Nous avions aussi vu, avec le bilan énergétique des vers endogés, comment un système de digestion mutualiste jamais décrit auparavant permettait à un ver – qui devait ingérer tous les jours jusqu’à 20 fois son poids de terre pour ne pas maigrir – de digérer 9% du 1,5% de matière organique que contenait ce sol, 95% de l´ énergie ainsi assimilée étant utilisée en activités mécaniques : creuser, ingérer et faire transiter toute cette terre en moins d´une heure à travers tout l’intestin (Lavelle, 1978). Ah ! Darwin et ses vers de terre, encore !! Ce faisant, les vers maintenaient 50% du volume de ce sol très pauvre, à peine pourvu de 5% d’une argile peu réactive, en macro-agrégats stables essentiels pour la séquestration du carbone, l´infiltration et le stockage de l´eau (Martin, 1990 ; Blanchart et al., 1997).
Nous mesurions enfin des effets positifs de l’activité endogée des vers de terre sur la croissance des plantes, parfois très importants. Ainsi, 72% des plantes testées lors de ces expériences ont répondu à l’activité des vers par un supplément de croissance (relativement aux plantes cultivées dans un sol privé de vers), de 56% en moyenne. Cinq mécanismes expliquent ces effets (Brown et al., 1999), dont en particulier : i) la stimulation des associations avec les microorganismes bénéfiques aux plantes (mycorhizes*et rhizobium*), ii) la production d´hormones de croissance et iii) la protection contre certains parasites, par la manipulation de l´expression des gènes de la plante (Blouin et al., 2005)
La théorie de l´auto-organisation nous a permis de concilier dans un même modèle la création d´habitats par les organismes via la formation de structures, agrégats ou pores, le travail mécanique impressionnant des organismes ingénieurs*, racines et invertébrés, et l´existence de relations mutualistes anisosymbiotiques* fortes entre macro et microorganismes du sol, permettant l´utilisation de ressources difficiles à digérer.
Ce modèle intègre aussi les relations antagonistes des réseaux trophiques et leur gestion des activités microbiennes dans les microsites où ils s´activent (Perry, 1995 ; Camazine et al., 2001). Tout ceci, dans une structure spatiale hiérarchique des sols, des micro-agrégats microbiens aux agrégats et pores de taille intermédiaire où vivent les microprédateurs, aux structures biogéniques millimétriques à pluri centimétriques des racines et des invertébrés ingénieurs*, aux catenas de sol et au paysage où se manifestent in fine les services écosystémiques, fruit de cette coopération multi-organismes et multi-échelles (Lavelle et al., 2016).

Figure 2 : Les surales, ilots émergés dans les savanes inondées de l´Orénoque colombien. Ces ilots sont construits par les vers de terre et les plantes, dans un processus d´auto-organisation Source : Zangerié et al., 2016, Plos One.
L’écologie du sol au service du futur : ingénierie biochimique et dynamique de l’auto-organisation
Comme la physique newtonienne fut remise en question par la physique quantique, quand l’échelle d’observation s’est affinée, l ‘écologie des traits biologiques, traits fonctionnels et traits d´histoire de vie, qui disjoint les fonctions des espèces en la série de traits qu´elles présentent (Lavorel & Garnier, 2002 ; Mouquet et al., 2015; De Bello et al., 2021) ou le quorum sensing qui crée des sauts qualitatifs dans la réponse des microorganismes aux stimulations externes (Miller & Blassner, 2001, et voir le Regard R85), explorent d´autres échelles et d´autres possibles. La connaissance de l’ingénierie biochimique qui accompagne les interactions entre plantes (Chave, et al., 2014), invertébrés et microorganismes n’en est qu’à ses débuts. Elle devrait permettre de mieux utiliser le processus d’auto-organisation dans la gestion et la restauration des sols et des écosystèmes.
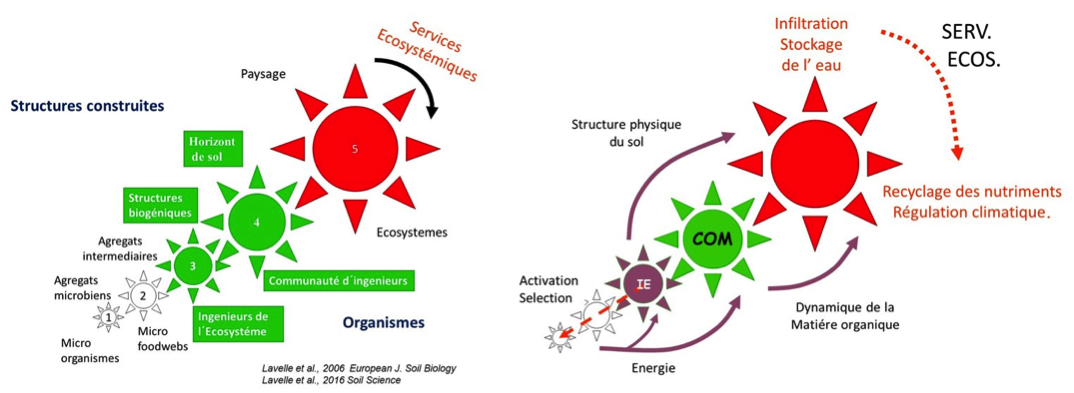
Figure 3 : le sol auto-organisé. A gauche : les organismes et les structures qu´ ils créent sont organisés en une suite d´échelles discrètes organisées de façon hiérarchique. Comme dans tout système hiérarchique, les mécanismes en jeu à une échelle donnée expliquent les propriétés émergentes observées à l´échelle supérieure, tandis que les processus actifs à cette échelle expliquent la configuration observée à l´échelle inférieure. Cette configuration permet l´existence de rétroactions entre les différents niveaux. A droite : Les ingénieurs de l´Ecosystème, macro invertébrés et racines, digèrent la matière organique du sol avec un système de digestion mutualiste en interaction avec les microorganismes du sol. Cette énergie est utilisée 1. Pour des actions physiques qui construisent les habitats du sol et permettent l’infiltration, le stockage et le transfert de l’eau ; 2. Pour transformer la matière organique morte en nutriments assimilables par les plantes (Recyclage des nutriments) ou la stocker sous des formes plus ou moins humifiées dans les agrégats de différentes tailles ou elle est physiquement protégée. Schéma P. Lavelle.
Il est important, par exemple, de comprendre pourquoi toutes les espèces de plantes ne s’associent pas à toutes les espèces de vers de terre dans le processus de consolidation des macro agrégats décrit par Zangerle et al. (2011). Important aussi de préciser la nature des relations entre la chaîne des microprédateurs et les organismes ingénieurs*. On connait le rôle essentiel des Protistes dans la boucle microbienne rhizosphérique* (Coleman, 1994 ; Bonkowski, 2004), mais les relations entre ces mêmes microprédateurs et les vers de terre ou les termites sont encore largement ignorées (Lavelle et al.,2004 ; Boyer et al.,2013).
On sait aussi que les microarthropodes sont plus nombreux et plus divers dans les sites où se concentrent les vers de terre (Loranger et al., 1998 ; Marinissen et Bok, 1988 ; Salmon, 2004 ; Grubert et al., 2016), mais une vision générale de ces relations fait cruellement défaut. Il faut aussi explorer le domaine et les possibilités immenses offertes par les plantes compagnes* et les microorganismes associés dans les cultures et dans la construction de paysages durables (qui n’impliquent pas seulement des légumineuses, cf. Chave et al., 2014 ; Blouin et al, 2015).
On pourra alors imaginer des modèles de simulation qui explorent la dynamique et les processus de l’auto-organisation, leurs relations avec toutes les formes de biodiversité, leurs aspects énergétiques, leurs liens avec les services écosystémiques, leur dynamique.
Cet agenda de recherche en appui à l’agroécologie et à l’écologie en général, doit remplir le vide cognitif et conceptuel dans lequel se perd souvent l’agriculture du futur. Permaculture, biodynamie, agroécologies inspirées de recettes éprouvées de leurs praticiens, mais certainement perfectibles et au moins vérifiables. De l’autre côté, celui de l’agriculture productiviste industrielle, cette vision pourrait – mais dans quelle mesure ? – amener à mieux respecter les lois incontournables de la durabilité, en l’extrayant du carcan idéologique et économique qui la soutient en dépit de tout, et en utilisant ses éléments utiles, dans un autre cadre.
Au-delà de cette étape qui permettra d’exploiter les champs ouverts par la reconnaissance du processus général d’auto-organisation, un autre grand chantier s’annonce, celui de la rénovation de la communication scientifique et l´élargissement du champ scientifique matériel à celui de la prise de conscience et de la spiritualité .
Une communication scientifique plus détendue
La grande crise environnementale actuelle consacre l´échec de la science à informer les citoyens de la gravité de la situation et à les motiver à ne pas accepter ce lent suicide collectif (Foucart, 2021). Les grands décideurs continuent à faire le minimum et les politiques publiques nécessaires pour lancer ce mouvement restent faibles. Il n´y a alors pas d´autre solution que d´alimenter un fort mouvement citoyen du bas vers le haut (bottom up, en anglais) qui permettra de parvenir à cette fin, dans un scenario de « mosaïque adaptative » proposé par le Millennium Ecosystem Assessment (2005).
La grande qualité des modèles et des prévisions publiés dans les rapports du Millennium Ecosystem Assessment et ceux du GIEC est certes un apport essentiel (MEA, 2005 ; IPCC, 2014 ; 2021). La crise (socio)écologique actuelle est une réalité anxiogène et la communication doit recourir à d´autres méthodes pour que le message ne parvienne pas seulement au cerveau et à la raison (Mouterde, 2021). Qu´il touche la sensibilité et donne à l´audience l’énergie et l´espoir nécessaire à la création urgente d´un autre monde.
Il faut à mon avis pour cela s´allier à l´art, et je peux témoigner que l´explication du changement climatique lié aux activités humaines se comprend mieux quand on met en scène une débonnaire planète Terre, en butte aux délires d´adolescents de sa dernière création, l´Homo autodéclaré sapiens, à l´ego démesuré. Patrick, l´exotique scientifique français dans le monde où je vis, lui explique avec cartes, tableaux et graphiques la nature de ses maux et le groupe musical Los Hijos de Gaia chante à sa manière le thème évoqué (https://www.youtube.com/watch?v=SkGRzeU6vKM&list=PLV1ae3tm89osDDUfpe3QVcZGJyoxg7v0d). La musique et la libre expression d´une chanson où tout peut être dit, l´incompréhension, la colère et la tristesse devant les dérives de toutes sortes d´une humanité perdue dans son délire egocentrique. On n´a jamais vu de tels sentiments exprimés dans des articles scientifiques. Le champ possible de cette rénovation pédagogique est sans limites. Suivant par exemple ces citoyens conscients qui relaient avec talent le message scientifique en y mettant humour, créativité … et colère (Gatineau, 2018, 2019).
Du dualisme au monisme ?
Descartes et les philosophes du siècle des lumières nous ont convaincus de séparer la sphère spirituelle de la sphère matérielle, passant d’une vision moniste unique du monde à la vision dualiste présente (Goswani et al., 1995). Cette révolution a permis l´avènement de l´ère moderne avec ses spectaculaires avancées technologiques et matérielles qui ont amélioré considérablement la vie matérielle de la plupart des humains tout en réduisant progressivement l´importance de la sphère spirituelle. Au bout de sa logique, cette percée a été si forte qu´elle a engendré la société matérialiste présente, son consumérisme compulsif comme objectif de vie et la perte de l´autre moitié de nous-mêmes, faite de conscience et d´esprit (Goswani et al., 1995). Les physiciens, passée la révolution quantique, sont désormais engagés dans la reconquête de cette unité.
L´écologue sensoriel perçoit cette dimension. La joie de découvrir les mille et une façons d´être une abeille ou un ver de terre, l’énergie puissante et douce qui émane d’une foret ancienne, le désespoir face à la laideur d´un sol détruit par l´abus des produits chimiques et des maltraitances mécaniques, la fascination de comprendre ce que font des organismes aussi peu dotés en structures cérébrales, avec aussi peu : la communication des arbres par les réseaux mycorhiziens, les débats chorégraphiques des abeilles, et, puisqu´il faut que Darwin la ramène encore une fois avec ses vers de terre, la fascinante histoire du ver de terre endogé mâle adulte qui est capable, à 40 cm de profondeur dans ce sol de savane, de repérer à plusieurs décimètres de distance, voire plus, dans le volume compact du sol, l´autre mâle adulte avec lequel il pourra s’ accoupler mais qu’il ne pourra rejoindre qu’une fois creusée la galerie qui les unira, dans un accouplement homosexuel mais néanmoins fécond (Lavelle, 1978).
Le dogme voulant que la conscience soit réservée à l´espèce humaine fait eau de toute part. Quelle est la conscience d’une plante ? d´une fourmi ? et comment pouvons-nous la percevoir ? Il nous appartient maintenant d´explorer ce potentiel et d´en faire le meilleur usage dans une gestion respectueuse des ressources naturelles. On pourrait penser que le type d’interaction inné qui dicte les relations d’un organisme avec les autres organismes et le milieu à son échelle, leur tient lieu de conscience, à l’instar des poissons d´un banc se situant par rapport aux voisins les plus proches ; mais alors, que penser de ce résultat de Martin (1990) qui montre -dans son modèle mécaniste DRILOTROP qui simule la dynamique des populations du ver de terre tropical Millsonia anomala et leur impact sur le sol – que si 5% au moins des individus ne se déplacent pas comme les autres en fonction des gradients d’ humidité, de température et de richesse nutritive du sol, la population finit par se trouver coincée dans une strate de sol défavorable à sa survie et disparait. Eloge de la différence ?
La méthode scientifique traditionnelle peut nous aider à progresser dans cette direction, mais sans les apports de la philosophie et de l´histoire et sans exploration de la spiritualité, ces progrès resteront limités. Mais par où commencer ? Décoloniser notre pensée écologique comme le suggèrent Trisos et al. (2021), nous reconnecter au vivant (Tassin, 2020) et réintégrer la vision des peuples premiers dans un modèle qui conserve les acquis de la modernité des lumières ? Ou pousser plus loin l´exploration de paradigmes plus « scientifiques » ? Toutes les approches peuvent s´avérer nécessaires, mais ce modèle est à réinventer… d´urgence.
Le mot de la fin, avec Darwin
En guise de conclusion, cette remarque de Darwin à la fin de son livre…
« Quand nous contemplons une vaste étendue couverte de gazon, nous devons nous rappeler que sa régularité, dont dépend une grande partie de sa beauté, est
principalement due au fait que toutes les inégalités ont été lentement nivelées par les vers de terre. C’est une sensation merveilleuse de réaliser que l’ensemble du sol superficiel sur une telle étendue est passé, et passera à nouveau après quelques années, par les intestins des vers. La charrue est l’une des plus anciennes et des plus importantes inventions humaines; mais bien longtemps avant qu´elle ait été inventée, la terre était déjà labourée, et continue de l´être, par les vers de terre. Il n´y a sans doute aucun animal qui ait joué un rôle aussi important dans l´histoire du monde que cette créature à l´organisation aussi simple. » (Traduction P. Lavelle)
Glossaire
- Anécique (ver de terre) : les vers de terre sont classés en trois groupes fonctionnels : les épigés qui vivent dans la litière et la compostent ; les endogés qui vivent dans le sol et consomment la terre et sa matière organique et les anéciques qui vivent dans des galeries sub verticales qui débouchent a la surface du sol ou ils vont récolter la litière qu´ils ingèrent mélangé au sol minéral. Les anéciques comprennent les plus grandes espèces qui peuvent mesurer jusqu´a 1m de long pour des espèces rencontrées au pays basque ou en Provence.
- Boucle microbienne rhizosphérique: boucle d´interaction, depuis la racine vers les bactéries, nématodes et protistes prédateurs de ces bactéries, avec un retour vers la racine. Elle permet à la racine en croissance de récupérer les nutriments extraits du sol par les bactéries qui se multiplient autour de la pointe racinaire, stimulées par la libération dans le sol d´exsudats racinaires riches en carbone assimilable. Ces bactéries, après avoir grossi et s´être multipliées, sont la proie de petits invertébrés, protistes ou nématodes dont les excrétions azotées sont absorbées par la racine.
- Chaîne trophique = réseau trophique: suite d´organismes qui, depuis les plantes et autres « producteurs primaires » -qui fixent le gaz carbonique de l´air et le transforment en structures carbonées, la biomasse végétale- consomment ces végétaux (consommateurs primaires), et sont eux-mêmes consommés par des prédateurs, (consommateurs secondaires, tertiaires etc…). L´énergie captée par les végétaux est ainsi transférée et accumulée vers des niveaux trophiques supérieurs. On considère qu´en moyenne, de l´ordre de 10% de la biomasse prélevée à un niveau (par ex. la biomasse végétale consommée par les herbivores) est accumulée dans la biomasse du niveau supérieur (ici, les herbivores). Les 90% restant entreront dans la chaine des décomposeurs, ensemble des organismes qui recyclent la matière morte, localisés pour leur plus grande partie dans les sols et les couches d´humus qui les recouvrent.
- Micorhizes : filaments de champignons attachés aux racines qui apportent à celle-ci l´eau et les nutriments qu´ils collectent en explorant un grand volume de sol et reçoivent en retour le carbone issu de la photosynthèse que la plante transporte vers les racines. On estime a 20% la proportion des produits de la photosynthèse ainsi apportés aux organismes du sol.
- Organismes ingénieurs des sols = ingénieurs de l’écosystéme = invertébrés ingénieurs des sols : organismes qui par leurs activités physiques et biochimiques – telles que creuser chambres et galeries, ingérer le sol et le faire transiter par leur tube digestif, mélanger la matière organique au sol, … – créent des habitats et autres ressources (OK ?) dans le sol pour d’autres organismes, avec un effet positif pour la plupart des organismes présents. Les principaux sont les vers de terre, les fourmis, les termites et les racines des plantes.
- Patron spatial: schéma de distribution spatiale des individus d´une espèce ou d´un ensemble d´espèces (communauté) dans l´espace représenté par un écosystème et son sol. Les vers de terre sont ainsi souvent distribués en taches plus ou moins bien individualisées, pas forcément superposées à des taches correspondantes de zones présentant de meilleures conditions de vie.
- Plantes compagnes: plantes associées aux cultures pour leurs effets bénéfiques sur la plante cultivée. Exemples : les légumineuses qui fixent l´azote de l´air, tel le trèfle, dans une prairie ; les piments qui attirent les nématodes parasites, protégeant les plants de maïs de leurs attaques.
- Relation anisosymbiotique: relation entre deux organismes de tailles très différentes (ex : ver de terre et bactérie).
- Rhizobium : genre de bactéries fixatrices de l´azote de l´air. Elles forment les nodules rosés que l´on peut observer sur les racines des plantes du groupe des Légumineuses (Famille des Fabacées) : trèfle, luzerne, féverole, haricots, lentilles…) apportant a celles-ci l´azote qu´elles nécessitent. La présence de légumineuses, en culture pure ou en co culture avec d´autres plantes qui ne bénéficient pas de cette association permet de se passer des engrais azotés d´origine industrielle.
- Rhizosphère : Voir ‘Boucle microbienne rhizosphérique’.
- Turricules: déjections de vers de terre qui déposées en surface peuvent avoir l´aspect d´une petite tour. Déposés pour l´essentiel à l´intérieur du sol (de l´ordre de 90% globalement pour tous les vers de terre) ils s´y accumulent, formant les grumeaux de sol appelés macro-agrégats par les physiciens du sol. Leur présence/accumulation donne au sol des propriétés physiques favorables à l’infiltration, le stockage et le transfert de l´eau de pluie, et à l´aération des racines. Ce sont aussi des sites très importants pour la conservation du carbone dans les sols, qui a ce titre sont considérés dans des modes de gestion qui favoriseraient le stockage du carbone avec des effets favorables pour le climat.
Annexe
Exercice proposé aux étudiants à leur entrée en maitrise :
Quelle théorie écologique permet d’interpréter la distribution de ces deux espèces de vers (dont l’effet sur la physique du sol est opposé), dans le schéma ci-dessous ?
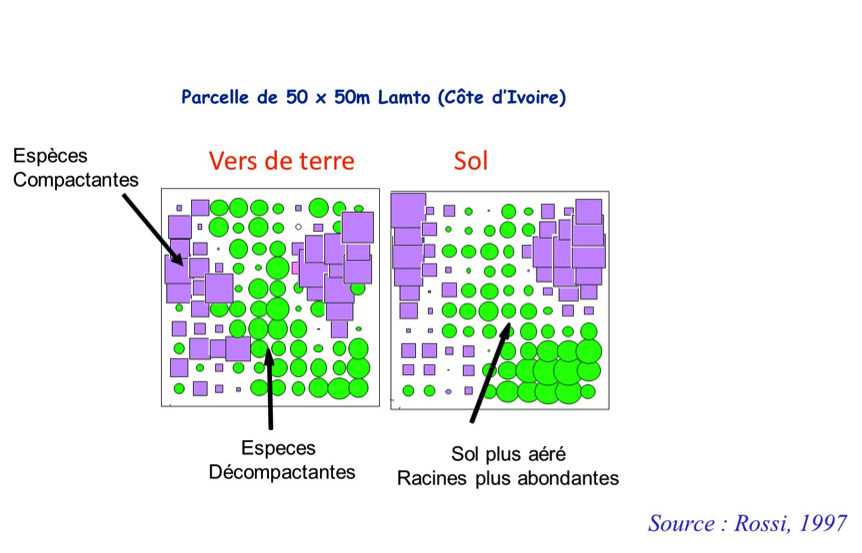
3 réponses sont possibles :
1. Exclusion compétitive
2. Réponse à des conditions de sol différentes
3. Mutualisme entre les deux groupes fonctionnels de vers de terre
Dans l’option 3, chaque groupe fonctionnel accumule progressivement dans le sol les structures qui serviront de nourriture á l’autre groupe. Les vers « compactants » ne reingèrent pas leurs turricules*, s´alimentant d’agrégats de sol de petite taille qui sont de fait les turricules* produits pas les espèces décompactantes dont la teneur en matière organique est élevée. Les espèces décompactantes ingèrent quant à elles les turricules formés/produits/ par les compactantes, y ajoutant de la matière organique qui provient probablement des racines fines mortes qui ont colonisé ces turricules ou des exsudats racinaires. On peut prédire que le patron spatial* observé s’inversera quand chaque groupe aura exploite complètement la tache où il est plus abondant. Une modélisation du processus montre la formation de cette distribution spatiale alternée (Barot et al., 2007). Des observations sur la dynamique des taches appuyant cette hypothèse font cependant défaut.
Remerciements
Merci à tous les étudiants en thèse, collègues chercheurs et agriculteurs de partout, avec qui nous avons exploré tant de sites, naturels ou non, expérimenté, discuté, partagé et progressé vers une vision à chaque fois plus vaste des sols et de leur gestion. Merci aussi à Anne Teyssèdre et Jérôme Cortet, éditeurs de ce Regard pour la SFE2, pour leurs nombreuses remarques, suggestions et corrections, ainsi qu’à la Société Française d´Écologie et Evolution elle-même pour favoriser cet espace de débats sans tabous ni restrictions.
Bibliographie
Barot S, Blouin M, Fontaine S, Jouquet P, Lata J-C. et al., 2007a. A Tale of Four Stories: Soil Ecology, Theory, Evolution and the Publication System. PLoS ONE 2(11)
Barot S., J. P. Rossi and P. Lavelle, 2007. Self-organization in a simple consumer-resource system, the example of earthworms. Soil Biology & Biochemistry 39 (9): 2230-2240.
Blanchart E., P. Lavelle, E. Braudeau, Y. Le Bissonnais and C. Valentin. 1997. « Regulation of soil structure by geophagous earthworm activities in humid savannas of Cote d’Ivoire. Soil Biology and Biochemistry 29 (3-4): 431-439.
Blouin M., Zuily-Fodil Y et al., 2005. Belowground organism activities affect plant aboveground phenotype, inducing plant tolerance to parasites. Ecology Letters 8 (2): 202-208.
Blouin M., Battle, K., Mathieu, J. & Lerch, T. 2015. Levels and limits in artificial selection of communities. Ecology Letters 18: 1040–1048
Bonkowski M., 2004. Protozoa and plant growth: the microbial loop in soil revisited. New Phytologist 162 (3): 617-631
Bottinelli N., Jouquet P. et al., 2015. Why is the influence of soil macrofauna on soil structure only considered by soil ecologists? Soil &Tillage Research. 146, 118–124.
Boucher D. H., S. James and K. H. Keeler. 1982. The ecology of mutualism. Ann. Rev. Ecol. Syst. 13. 315-347.
Boyer, J., G. Reversat, P. Lavelle and A. Chabanne. 2013. Interactions between earthworms, and plant-parasitic nematodes. European Journal of Soil Biology 59: 43-47.
Camazine S., J. L. Deneubourg et al, 2003. Self-organization in biological systems, Princeton University Press. 538p.
Chave M., M. Tchamitchian and H. Ozier-Lafontaine, 2014. Agroecological Engineering to Biocontrol Soil Pests for Crop Health. Sustainable Agriculture Reviews, H. Ozier-Lafontaine and M. Lesueur-Jannoyer, Springer International Publishing. 14: 269-297.
Coleman D. C. 1994. The microbial loop concept as used in terrestrial Soil Ecology studies. Microbial Ecology 28 (2): 245-250.
Darwin C. 1881. The formation of vegetable mould through the action of worms with observations on their habits. London, Murray.
De Bello F., S. Lavorel et al., 2021. Functional trait effects on ecosystem stability: assembling the jigsaw puzzle. Trends in Ecology & Evolution 36 (9): 822-836.
Descola P., 2005. Par-delà la nature et culture, Paris, Gallimard, Bibliothèque des sciences humaines.
Erktan, A., M. C. Rillig, A. Carminati, A. Jousset and S. Scheu. 2020. Protists and collembolans alter microbial community composition, C dynamics and soil aggregation in simplified consumer-prey systems. Biogeosciences 17 (20): 4961-4980.
FAO, ITPS, GSBI, SCBD, and EC. 2020. State of knowledge of soil biodiversity – Status, challenges and potentialities, Report 2020. Rome, FAO. https://doi.org/10.4060/cb1928en
Filser J., J. H. Faber et al., 2016. Soil fauna: key to new carbon models. Soil 2 (4): 565-582.
Foucart S. 2021. Le “biodiversité-scepticisme”, plus discret que celui contre le dérèglement climatique, est en un sens bien plus inquiétant. Le Monde, 25/05/2021.
Gatineau C., 2018. Eloge du ver de terre. Flammarion, Paris.
Gatineau C. & Corré S., 2019. Eloge de l’abeille. Flammarion, Paris 245p.
Goswani A., Reed R.E. and Goswani M., 1995. The self-aware universe. How consciousness creates the material world. Tarcher Penguin, New York, 317p.
Grubert D., O. Butenschoen M. Maraun and S. Scheu,. 2016. Understanding earthworm – Collembola interactions and their importance for ecosystem processes needs consideration of species identity. European Journal of Soil Biology 77: 60-67.
Gunderson L. H. & Holling C.S. (Eds.), 2001. Panarchy. Understanding transformations in human and natural systems. Island Press.
IPCC, 2004. ARC synthesis report 2014. https://www.ipcc.ch/report/ar5/syr/ .
IPCC, 2021. AR6 Climate Change 2021 – Sixth Assessment Report – IPCChttps://www.ipcc.ch/report/ar6/wg1/
Jagers op Akkerhuis, G. 2015. Evolution and transitions in complexity. The science of hierarchical organization on nature. Springer, 295p.
James S. W., C. Csuzdi et al., 2021. Comment on “Global distribution of earthworm diversity”. Science 371: 6525.
Kropotkin P., 1902. Mutual aid: A factor in evolution. London, Heineman.
Lavelle P., 1978. Les Vers de Terre de la savane de Lamto (Côte d’Ivoire): peuplements, populations et fonctions dans l’écosystème, Paris VI. Publication du Laboratoire de Zoologie de l’ENS.. 301.
Lavelle P., 2000. Ecological challenges for soil science. Soil Science 165 (1): 73-86.
Lavelle P. & A. V. Spain. 2001. Soil Ecology. Amsterdam, Kluwer Scientific Publications. 654p.
Lavelle, P., M. Blouin et al., 2004. Plant parasite control and soil fauna diversity. Comptes Rendus Biologies 327 (7): 629-638.
Lavelle P., A. Spain et al., 2016. Ecosystem Engineers in a Self-organized Soil: A Review of Concepts and Future Research Questions. Soil Science 181 (3-4): 91-109.
Loranger G., J. F. Ponge, E. Blanchart and P. Lavelle., 1998. Impact of earthworms on the diversity of microarthropods in a vertisol (Martinique). Biol Fertil Soils 27: 21-26.
Marinissen J. C. Y. and J. Bok, 1988. Earthworm amended soil structure. Its influence on collembola populations in grassland. Pedobiologia 32 (3-4): 243-252.
Martin S., 1990. Modélisation de la dynamique et du rôle d’une population de vers de terre Millsonia Anomala dans les savanes humides de Côte d’Ivoire. Université Pierre et Marie Curie. Paris VI. 205p.
MEA, 2005. Millennium Ecosystem Assessment. https://www.millenniumassessment.org
Miller M.B. and Nassler B.L., 2001. Quorum sensing in bacteria. Annual Review of Microbiology 55, 165-199.
Mouquet, N., Y. Lagadeuc, V. Devictor et al., 2015. « REVIEW: Predictive ecology in a changing world. » Journal of Applied Ecology 52 (5): 1293-1310.
Mouterde P. 2021. Il faut amener chacun à reprendre contact avec le vivant. Le Monde, 2/09/21.
Nielsen U. N., D. H. Wall and J. Six. 2015. Soil Biodiversity and the Environment. Annual Review of Environment and Resources 40: 63-90.
Orgiazzi A., Singh B., Wall D., 2015. Global Soil Biodiversity Atlas, Publications Office of the European Union, Luxembourg, 2015. ISBN 978-92-79-48168-0 (online),978-92-79-48169-7 (print),978-92-79-74163-0 (Kindle), JRC95584
Perry D. A., 1995. Self-organizing systems across scales. TREE 10: 241-245.
Price P W, 1984. Alternative paradigms in Community Ecology. In A New Ecology : Novel Approaches to interactive systems. P W Price, C N Slobodchikoff and W S Gaud. New York, John Wiley: 1-14.
Rossi J. P., 1997. Rôle fonctionnel de la distribution spatiale des vers de terre dans une savane humide de Côte d’Ivoire. Ecology. Paris, Paris VI: 298.
Salmon S., 2004. The impact of earthworms on the abundance of Collembola: improvement of food resources or of habitat? Biology and Fertility of Soils 40 (5): 323-333.
Thakur M. P., H. R. P. Phillips, U. Brose et al., 2020. Towards an integrative understanding of soil biodiversity. Biological Reviews 95 (2): 350-364.
Trigo D, Martin A and Lavelle P., 1992. A mutualist system of digestion in temperate earthworms, Allolobophora molleri and Octolasium lacteum. Act. Zool. Fenn. 196.
Trisos, C.H., Auerbach, J. & Madhusudan K., 2021. Decoloniality and anti-oppressive practices for a more ethical ecology. Nature,Ecology and Evolution. https://doi.org/10.1038/s41559-021-01460-w
Volterra, 1926. Fluctuations in the abundance of a species considered mathematically, Nature 118: 558-60.
Zangerle A., A. Pando and P. Lavelle, 2011. Do earthworms and roots cooperate to build soil macroaggregates? A microcosm experiment. Geoderma 167-68: 303-309.
Zangerlé A., D. Renard, J. Iriarte, S. L.E., A. K.L, J. Juilleret and D. Mc Key. 2016. The Surales, self-organized earth-mound landscapes made by earthworms in a seasonal tropical wetland. PLoS One 11 (5): e0154269.
Regards connexes :
Regards sur les sols : https://sfecologie.org/tag/sols/
Regards sur les interactions : https://sfecologie.org/tag/interactions/
Regards sur le fonctionnement des écosystèmes : https://sfecologie.org/tag/fonctionnement/
Sur l’agriculture : https://sfecologie.org/tag/agriculture/
Sur les mécanismes : https://sfecologie.org/tag/mecanismes/
Sur gestion et gouvernance : https://sfecologie.org/tag/gestion-et-gouvernance/
—–
Edition : Anne Teyssèdre et Jérôme Cortet, mise en ligne: A. Teyssèdre.
——-
Bonjour,
Ma question : pourquoi ce regard frais de Lavelle n’est-il pas édité sur papier afin de lui donner un peu d’éternité pour qu’il soit lu, vu et senti par les générations futures ?
Merci Patrick pour cette vue éclairée à une époque où beaucoup trop avancent à tâtons, phares éteints. Et bien entendu qu’il faut appendre à se décoloniser en permanence, c’était d’ailleurs l’un des secrets des sorciers yaquis du nord Mexique.
Entièrement d’accord avec Christophe Gatineau. On aimerait vraiment voir ce regard publié sous une forme stable, destinée à être encore visible dans les décennies, voire les siècles à venir. Un vent d’air frais dans l’écologie, et pas seulement du sol, un « coup de torchon » aussi, que n’aurait pas renié un Bertrand Tavernier. Merci Patrick pour ce regard salutaire, empreint d’une profonde culture et d’une entière sincérité…
Patrick Lavelle a raison de mentionner « l ́échec de la science à informer les citoyens de la gravité de la situation ». Le problème est l’écologie (la science) et non l’écologisme (sa transcription dans le monde politique). Qu’EELV fasse moins de 5% aux présidentielles 2022 exactement quand les effets de la crise écologique commencent à se faire ressentir pour de vrai par l’ensemble des citoyens est révélateur : Ce n’est pas de sa faute, EELV fait ce qu’il peut avec l’écologie qu’on lui donne. C’est cette écologie qui est en dessous de 5%.
Mais en lisant Patrick Lavelle, je doute que sa proposition de revenir à un monisme soit la bonne. Pour beaucoup de petites raisons (ce serait une régression, l’idée du bon sauvage a été prouvée incohérente, si spiritualité il y a alors ce supposé monisme est un dualisme, etc.), et pour une grande raison : ce papier de Lavelle est très dualiste, pourquoi alors vouloir un monisme ? Ce papier oppose vivant à non vivant à bas bruit, alors qu’il y a urgence à crier ce dualisme.
Il y a donc beaucoup plus simple que de lancer une polémique « décolonisation de nos pensées » ou une « exploration de paradigmes plus scientifiques ». Il suffit à l’écologie occidentale d’assumer sa science. Le dualisme existe. Il n’oppose pas l’esprit à la matière mais la matière avec esprit (le vivant) à la matière sans esprit (l’inerte). Quelques part dans l’histoire, un cristal muni du pouvoir de se reproduire avec mutation s’est de ce fait retourné contre le reste de la matière. On n’a même pas besoin de surcharger l’écologie avec la spiritualité. Elle peut bien rester une science matérialiste en laissant le problème dur de la conscience à la philosophie de l’esprit (qui par ailleurs s’en occupe très bien). Il suffit juste de reconnaître que l’esprit ou la conscience sont une des résultantes de l’intelligence inhérente à la matière vivante, qui est celle de se reproduire avec mutation, et donc de pouvoir assimiler son contexte autant mécaniquement que métaphoriquement.
Si l’écologie arrivait à faire passer en elle puis à la société le message que la vie lutte (travaille) contre l’inerte, que cela est une vérité comme l’est la rotondité de la Terre, et non une force (l’écologie n’est pas du vitalisme) ou une règle (l’écologie n’est pas de la politique) alors cela est suffisant pour sortir de la crise. Le banquier comprend avec cela la situation, le politique aussi, etc.
Merci Patrick pour cette vision rafraîchissante de l’écologie et de la vie… En effet, il serait bon de chercher un autre medium pour diffuser ces idées! Je suis assez horrifié par la rélégation du mythe du bon sauvage qui semble faire très plaisir au mâle blanc qui peut-être malgré lui continue à se sentir supérieur. Le sauvage n’est pas « bon » mais il est probablement plus apte à vivre en harmonie avec son environnement que le « bon » civilisé avec toute sa science pédante et surtout une arme de destruction massive de la biosphère et de notre spiritualité. Je conseille à ce titre le très intéressant (et très insuffisant) « Contre la méthode » de Paul Feyerabend pour remettre la science à sa place et l’empécher de nous détruire complètement.
La physique quantique a montré il y a quelques décennie que la matière n’est qu’une forme particulière d’énergie, la masse de nos particules n’étant qu’une convention de calcul. Un tel résultat devrait nous aider à ne plus séparer mais à transcender notre matérialisme et notre attachement maladif aux choses qui sont probablement une manisfestation de notre peur-panique de la mort. À ce titre, les sauvages sont souvent bien plus pragmatiques et courageux face à la mort que nous (les blancs cartésiens ou nous).
Pour terminer en citant Darwin (c’est très chic chez les écolos de planton), « ça n’est pas le plus fort ni le plus intelligent qui survit, c’est celui qui est le plus disposé à s’adapter ». Dans un scénario (celui du dernier rapport de l’IPCC) où un réchauffement de 4˚C devrait causer la disparition de 95% de la biodiversité et 90% de la population humaine, qui croyez-vous aura le plus de chance de survie: le cartésien avec sa science toute puissante qui buge quand la connection internet fait défaut où le « bon » sauvage avec son arc et ses flèches? Profitons bien de la vie car la prochaine phase du jeu s’intitule: Survival. Bonne chance à touTEs!
Bonjour Guillaume,
Je suppose que la pique du mâle blanc se sentant supérieur au bon sauvage s’adresse à moi puisque c’est moi qui ai introduit cette expression dans le fil de discussion. Je m’excuse alors, je me suis mal fait comprendre.
Je n’ai rien contre les autres civilisations que celle du mâle blanc au contraire je désire les protéger (et une part de moi fait partie d’une de celle-ci). Car non, les gars avec les arcs et les flèches ne survivront pas. Ils disparaîtront bien avant les autres. Ils ne pourront rien face au changement climatique, au changement sanitaire ou au changement écologique etc qui ravagent leurs terres et rien non plus contre les mouvements survivalistes armées organisées en bande ou même carrément en état voyou. Ce truc des indigènes mieux placés pour survivre est encore un mythe. Il suffit de voir en ce moment même ce qui leurs arrivent.
Si j’ai proposé de ne pas chercher à s’inspirer des sciences autres qu’occidentales pour corriger la science occidentale, c’est que cela est le chemin normal de la science. C’est le chemin normal et aussi le plus robuste. C’est à notre écologie occidentale de convaincre notre philosophie occidentale que la vie lutte contre l’inerte pour de vrai et non métaphoriquement.
Je m’excuse encore de vous avoir horrifié. Je m’étais mal exprimé.