
La Société Française d’Ecologie (SFE) vous propose le regard de Emmanuel Gheerbrant sur la diversification des mammifères.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
Origine et diversification des mammifères modernes
Emmanuel Gheerbrant
CNRS-MNHN, UMR-CNRS 7207, CR2P – Centre de Recherches sur la Paléobiodiversité et les Paléoenvironnements
( Fichier PDF )
Regard R5, édité par Anne Teyssèdre
———
Mots clés : Phylogénie, mammifères, évolution, biodiversité, espèces, Tertiaire, paléontologie.
———
Dans la diversité des vertébrés actuels, les mammifères occupent une place majeure. Pas seulement en raison de notre place parmi eux, ni de leur diversité spécifique (4500 espèces) bien inférieure à celle des poissons osseux (50 000 espèces) ou des oiseaux (9700 espèces). Les mammifères actuels se singularisent avant tout par leur spectaculaire diversité adaptative. De forme et de taille remarquablement variées, ils ont colonisé de nombreux milieux, sur terre, dans la terre, dans les arbres, les grottes, la mer, les airs, les déserts, les montagnes, etc… Le plus important groupe actuel est celui des placentaires, auquel nous appartenons. Il comprend 4200 espèces actuelles, de la minuscule musaraigne nocturne aux formidables éléphants et aux baleines, en passant par les lions et les singes. A côté des placentaires, les marsupiaux («mammifères à poche ») comptent 300 espèces actuelles, et les monotrèmes (mammifères qui pondent des œufs) quelques unes, dont l’ornithorynque.
Origine et débuts de l’histoire des mammifères

Jeholodens : petit mammifère triconodonte du Crétacé (Chine, 125 Ma, Ji et al. 1999)
L’histoire des mammifères remonte comme celle des dinosaures au Trias, il y a 220 millions d’années (Ma). Celle des mammifères modernes, marsupiaux et placentaires, est bien plus tardive ; leurs plus anciens fossiles tels que Sinodelphys et Eomaia datent du début du Crétacé, il y a 130 Ma.
Pendant l’ère Secondaire ou Mésozoïque (220-65 Ma), aux temps des grands dinosaures terrestres, prédominent surtout des lignées archaïques de faible diversité morphologique et adaptative représentées par de petites espèces insectivores ou carnassières, arboricoles ou terrestres. Un grand groupe d’herbivores éteint dominait à leurs côtés, les multituberculés, à l’allure proche de celle des rongeurs. De récentes découvertes d’espèces aux adaptations inattendues, auparavant seulement connues pendant l’ère Tertiaire ou Cénozoïque (65-0 Ma), ont toutefois montré que le monde des mammifères de l’époque des dinosaures était beaucoup moins uniforme que ce que l’on pensait. Elles sont figurées par des fossiles exceptionnels conservant non pas seulement les dents, mais la plus grande partie du squelette, et parfois même des empreintes de parties moles telles que la fourrure ou l’estomac et son contenu.
Une de ces espèces du genre Repenomamus, vivant au Crétacé inférieur (125 Ma) en Chine, était un grand mammifère prédateur de la taille d’un chien (15 kg) qui se nourrissait de petits dinosaures. Fruitafossor du Jurassique (150 Ma) des Etat-Unis avait des dents de tatous et des mœurs fouisseuses comme les taupes. Volaticotherium (15 cm), du même gisement que Repenomamus, était comme l’écureuil volant capable de vol plané entre les arbres. Castoracaudata, long de 40 cm, du Jurassique (164 ma) de Mongolie savait bien nager et mangeait des poissons.
Les premiers représentants des mammifères modernes (marsupiaux, placentaires, et leurs proches ancêtres), connus au début du Crétacé, marquent l’apparition de la mastication vraie, avec l’invention de la molaire moderne dite « tribosphénique » dont la couronne, munie de trois pointes ou cuspides, est capable de couper, percer et surtout broyer la nourriture. C’est cette nouvelle fonction-clef de broyage des dents qui va favoriser l’évolution et l’explosion des espèces omnivores et herbivores, une niche écologique restée auparavant inexploitée par les mammifères primitifs.
La radiation des mammifères modernes
Si l’origine des marsupiaux et placentaires est commune, et probablement située en Asie, leurs radiations sont indépendantes. Au plan géographique, les marsupiaux se sont diversifiés dans la ‘guirlande’ Amériques Nord et Sud-Antarctique-Australie, tandis que les placentaires se sont épanouis dans l’ensemble de l’Ancien Monde et de l’Amérique du Nord. Les deux radiations montrent nombre de convergences, parfois spectaculaires.
Les archives fossiles indiquent que l’explosion des groupes modernes ne commence pas avant le Tertiaire, même si quelques groupes sans descendants se sont diversifiés au Crétacé. Elle est très probablement en relation directe avec la conquête des niches laissées vacantes après l’extinction des dinosaures terrestres (non aviens).

Eomaia, plus ancien représentant des mammifères euthériens (Chine, 125 Ma, (Ji et al., 2002)
Si les événements de la transition Crétacé-Tertiaire (65 Ma) ont profité aux mammifères modernes, ce passage n’a pas été sans douleur pour d’autres groupes. A l’exception des multituberculés et des monotrèmes, tous les mammifères primitifs non « tribosphéniques » disparaissent à cette époque (Fig.). Même les mammifères tribosphéniques connaissent d’importantes extinctions. Jusqu’à 90 % des espèces de marsupiaux disparaissent ainsi en Amérique du Nord à la fin du Crétacé.
La radiation des marsupiaux est relativement localisée. Quelques lignées, aujourd’hui pour la plupart éteintes, se sont diversifiées en Amérique du Nord et du Sud. Mais leur principal centre de radiation moderne correspond à l’Australie, où ils sont aujourd’hui les plus diversifiés. Le monde des marsupiaux australiens illustre un exemple typique de radiation adaptative endémique, avec de nombreuses formes aux adaptations variées (kangourou, koala, loup marsupial), certaines atteignant la taille d’un rhinocéros (Diprotodon, 1,8 Ma); ses plus anciens jalons fossiles remontent au début de l’Eocène (55 Ma).
Les placentaires franchissent sans encombre la transition Crétacé-Tertiaire, mais leur histoire lors de ce passage reste mal connue, documentée de façon très localisée, surtout en Amérique du Nord et en Asie. D’après les fossiles, la radiation des placentaires commence au début du Tertiaire. Elle atteint son apogée à l’Eocène où la majorité des ordres modernes apparaissent, peut être en liaison avec le grand réchauffement climatique alors observé. Cette période-clef d’évolution est caractérisée par un accroissement très rapide de la taille des espèces et de nombreuses innovations morphologiques liées à de nouvelles adaptations alimentaires et locomotrices.
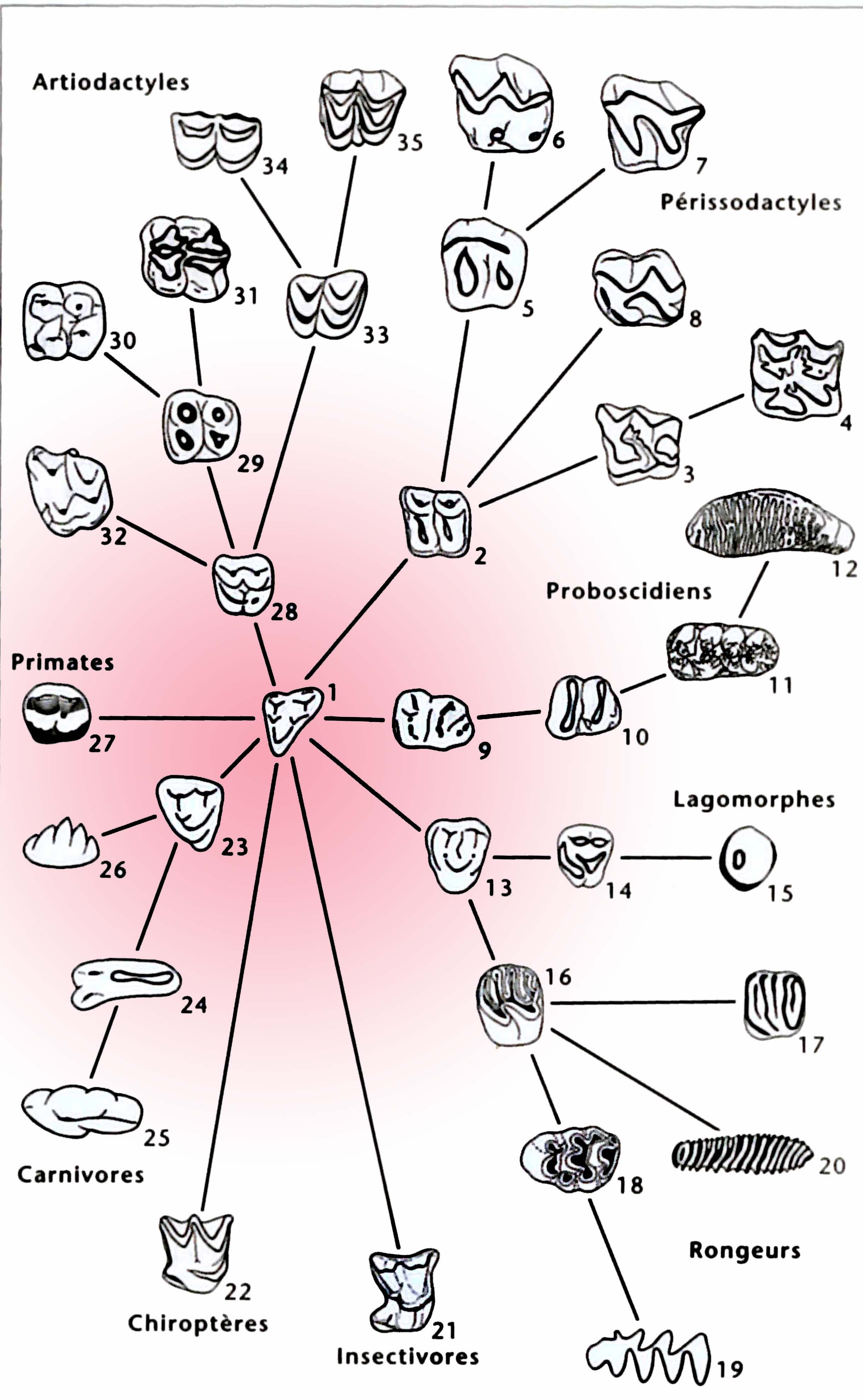
« Arbre à dents » des mammifères placentaires
(Hartenberger, 1992)
L’ensemble des grands plans adaptatifs des mammifères actuels sont développés à l’Eocène, vers –55 Ma : fouisseurs, insectivores, carnassiers, omnivores et herbivores spécialisés (folivores, brouteurs, etc..). C’est ainsi qu’apparaissent parmi les groupes modernes les premiers artiodactyles (ruminants, cochons…) et leurs proches cousins cétacés (baleines), périssodactyles (chevaux, rhinocéros), chiroptères (chauves-souris), carnivores (lions, chiens), lagomorphes (lapins), siréniens (lamantins), primates lémuriformes (lémurs) et simiiformes (singes).
Au Paléocène (-65 à -55 Ma), seuls de rares ordres modernes de placentaires sont représentés, et encore de façon très fragmentaire : xenarthres, insectivores (des familles éteintes), proboscidiens, primates et rongeurs. La plupart des groupes apparus au Paléocène, dont les fameux « condylarthres », appartiennent en fait à des lignées primitives qui vont s’éteindre rapidement. Ce cliché de l’explosion des groupes modernes à l’Eocène est cependant probablement exagéré par les lacunes fossiles du Paléocène.
Au cours de l’Eocène (-55 à -34 Ma) de nombreuses lignées évoluent de façon endémique et les diversités se stabilisent. A l’Oligocène, entre – 34 et – 23 Ma, la plupart des familles modernes de placentaires sont établies, et le groupe des singes de l’Ancien Monde et de l’Homme (catarrhiniens) apparaît en Afrique.
Pour en savoir plus
Sagascience Evolution, dossier « Radiation des mammifères »
Exposition MNHN, Dans l’ombre des Dinosaures
Livres :
Y.Coppens, K. Padian, A.de Ricqles, P. Taquet (eds)(2009). Histoire évolutive de la Vie. Comptes Rendus Palevol (Elsevier Masson), Volume 8, n° 2-3.
J.F. Eisenberg (1981). The Mammalian Radiations. University Chicago Press, Chicago. 610 p.
J.-L. Hartenberger (2001). Une brève histoire des mammifères, bréviaire de mammalogie. Belin / Pour la Science. 288 p.
Z. Kielan-Jaworowska, R. Cifelli, Z. Luo (2004). Mammals from the age of dinosaurs : origins, evolution, and structure. New York: Columbia University Press. 631 p.
D. R. Prothero (2006). After the Dinosaurs. The Age of Mammals. Indiana University Press. 362 p.
K.D. Rose (2006). The Beginning of the Age of Mammals. Johns Hopkins University Press, Baltimore. 428 p.
K.D. Rose & J. D. Archibald (2005). The Rise of Placental Mammals. Origin and Relationships of Major Extant Clades. The Johns Hopkins University Press. 260 p.
La diversification des mammifères au Secondaire et leur radiation au Tertiaire soulève de nombreuses questions écologiques et évolutionnistes, parmi lesquelles :
– Globalement, pourquoi une telle diversification écologique des mammifères comparée à celles d’autres lignées de vertébrés tétrapodes apparues vers la même époque (230 Ma), tels que les dinosaures (terrestres + oiseaux) en particulier ?
– Ou, pour se concentrer sur le Tertiaire : pourquoi les mammifères modernes (placentaires et marsupiaux) ont-ils exploré et envahi nettement plus de niches écologiques que les oiseaux modernes ?
– Pourquoi les mammifères modernes et les oiseaux modernes ont-ils surmonté la crise géologique et écologique de la fin du Crétacé il y a 65 Ma, alors que les grands dinosaures terrestres et la plupart des mammifères ‘primitifs’ n’ont pu le faire ?
(Quels étaient les facteurs de résilience/vulnérabilité : taille, âge à maturité, fécondité, régime alimentaire, mobilité, plasticité comportementale…)
– Pourquoi les oiseaux modernes ont-ils laissé des mammifères nouveaux-venus, chauves-souris en devenir/évolution, occuper la niche écologique des insectivores aériens nocturnes ?
(Problème de la vision nocturne chez les oiseaux, peu de lignées d’espèces nocturnes…)
– Que donne la confrontation contemporaine entre marsupiaux et placentaires – hommes et espèces introduites – en Australie, depuis 40.000 ans ?
(Décimation de la mégafaune australienne il y a 40.000 ans, confrontation actuelle)
– Quels enseignements tirer de l’évolution passée des mammifères (diversification, extinctions, facteurs de résilience et de vulnérabilité des populations…) pour comprendre, anticiper et éventuellement freiner l’érosion actuelle des mammifères, face aux changements globaux ?
Pour Répondre à Anne (Teyssèdre) : La radiation des mammifères : « Pourquoi, comment, pourquoi eux ? »
Tout d’abord, il faut souligner que la radiation adaptative des mammifères modernes est un phénomène fondamentalement opportuniste, correspondant à la conquête de niches écologiques libérées à la suite des extinctions massives de la fin du Crétacé. Les découvertes fossiles montrent de fait de nombreuses convergences et le rôle important des contraintes fonctionnelles et structurales dans les mécanismes et modes évolutifs de cette radiation.
La compréhension de la biologie et de l’écologie de la radiation adaptative des mammifères modernes est nécessairement actualiste (comparaison avec les espèces actuelles) et indirecte, fondée sur les fossiles (parties dures du corps, squelette).
Elle fait l’objet de nombreux travaux qui vont des analyses moléculaires et génétiques, à l’embryologie, l’ « évo-dévo », la recherche de terrain (fouilles), la biochronologie des faunes fossiles, l’anatomie comparée et fonctionnelles, et bien sûr la systématique et la phylogénie, sans oublier l’étude du cadre paléoenvironnemental de la transition Crétacé-Tertiaire (KT).
Plusieurs facteurs- clefs de cette radiation sont identifiés à l’issue de ces études.
1. Physiologie, métabolisme
Il n’y a sans doute rien de plus important dans la vie des mammifères que leur métabolisme d’animaux à « sang chaud » et température constante, ce que l’on appelle endothermie et homéothermie. C’est une grande différence physiologique avec la plupart des autres vertébrés, exception faite des oiseaux. L’endothermie favorise les réactions enzymatiques et la biochimie cellulaire, et permet un métabolisme élevé. Le métabolisme de base des mammifères est 6-10 fois plus élevé que chez les reptiles ectothermes, et il est 10-15 fois plus élevé en pleine activité qu’au repos. L’homéothermie permet à cette machinerie physiologique et biologique « dopée » de tourner de façon optimale et constante, indépendamment des conditions extérieures.
Endothermie et homéothermie sont indubitablement à la base du succès adaptatif et évolutif des mammifères. Etablies très tôt, elles leurs ont permis de s’adapter à des environnements climatiques et géographiques très variés – de l’équateur aux pôles, et des montagnes aux profondeurs océaniques – et également aux niches nocturnes. Les mammifères sont de fait plus actifs que les reptiles ectothermes, et notamment dans des conditions de basses températures. Cela a probablement favorisé la niche refuge nocturne (et insectivore) des mammifères anciens du Crétacé.
De la même façon, le métabolisme élevé, la structure de l’oreille moyenne perfectionnée (ex. chaine des osselets) chez les mammifères, qui a permis le développement de l’écholocation, et un mode de vie ancestral arboricole, pourraient expliquer pourquoi les mammifères de l’ordre des chauves-souris ont pu coloniser la niche insectivore nocturne, à la différence des oiseaux.
2. Denture et régime alimentaire
Un des inconvénients majeurs du métabolisme élevé des mammifères est le grand coût énergétique. 80 % de la nourriture des mammifères est ainsi consacré au maintien de la température élevée ; par comparaison, les reptiles ectothermes tels que les crocodiles consomment proportionnellement 10 fois moins de nourriture.
Pour pallier à ces grands besoins énergétiques, les mammifères ont inventé une denture perfectionnée particulièrement efficiente et, chez les deux groupes modernes de thériens marsupiaux et placentaires, la mastication. Cette denture se caractérise par une spécialisation des dents en fonction de leur position (ICPM), la réduction du nombre de générations dentaires (une denture de lait et une denture définitive) qui a permis un emboitement (occlusion) précis et complexe des dents supérieures et inférieures. Les thériens modernes ont en outre innovés avec la molaire dite « tribosphénique » capable comme les autres groupes de percer et couper, mais capable aussi de broyer. Avec cette dent multi-usage et des mouvements de mastication non plus seulement verticaux mais également latéraux, ils ont inventé la mastication qui permet de broyer plus finement la nourriture, par différence avec les reptiles qui avalent sans mâcher et digèrent de gros morceaux, parfois des individus entiers (penser aux serpents notamment)…
Grace à cette denture perfectionnée et à la mastication, les mammifères tirent un plus grand profit et rendement de la nourriture, une carte de plus à leur adaptabilité. Cette denture qui était très généralisée chez les formes ancestrales omnivores et insectivores du Crétacé, a évolué de façon spectaculaire avec la conquête du monde au Tertiaire; elle s’est diversifiée et spécialisée dans sa morphologie et sa fonction dans une multitude de niches alimentaires (plancton, poissons, racines, herbes, nectar, chaire, coquillages, fourmis, etc..), correspondant à autant de stratégies trophiques…. Le perfectionnement de l’appareil dentaire a en particulier permis la colonisation de la niche herbivore et l’explosion la plus spectaculaire des lignées chez les placentaires (petits primates frugivores comme le loris, grands brouteurs comme les vaches et chevaux, opportunistes comme les cochons et singes, mangeurs d’algues comme les siréniens, etc..).
3. Reproduction
Le mode de reproduction des mammifères est un autre élément notable de leur succès, notamment par rapport aux reptiles. La viviparité des mammifères modernes (thériens) procure une plus grande indépendance du milieu. Les portées (4 à 8 petits) sont plus petites et la mortalité infantile est plus faible que chez les reptiles, par suite d’investissement et soins parentaux (ex. allaitement, typiquement mammalien) plus importants. Les générations sont également plus courtes, spécialement chez les petits mammifères, prédominants au Crétacé. Il en résulte une évolution plus rapide, et une plus grande adaptabilité génétique (importance/dynamisme du génome). C’est probablement un des facteurs de la résistance du groupe aux événements de la limite KT.
4. Locomotion
Le mode de locomotion des mammifères du Crétacé était de type quadrupède, terrestre ou arboricole. Cette locomotion « généraliste » leur a permis d’occuper des habitats variés et de nombreux territoires géographiques. Sur ce plan les oiseaux étaient plus spécialisés, avec des membres adaptés au vol à l’avant et à la course à l’arrière. La locomotion des mammifères a évolué rapidement au début du Tertiaire, de concert avec les nouvelles stratégies trophiques, dans diverses spécialisations remarquables illustrées par des espèces fouisseuses (taupes), nageuses (cétacés), coureuses (chevaux, artiodactyles), sauteuses (lapins), grimpeuses (primates), et volantes (chauves-souris).
Les mammifères volants et arboricoles ont des taux de mortalité plus faibles et des durées de vie plus longues, en raison notamment d’une pression de prédation moins forte. C’est particulièrement le cas des primates. Les avantages adaptatifs sont significatifs, particulièrement pour les petites espèces. C’est une autre explication de l’importance de la niche arboricole refuge chez les mammifères du Crétacé, et c’est probablement aussi en lien avec la radiation florissante des oiseaux et des chauves -souris.
5. Taille
La petite taille des mammifères ancestraux du Crétacé (< 1kg, plus fréquemment 200 à 5 g) est classiquement reliée à la compétition avec les lignées de reptiles contemporains, outre leur régime insectivore. Celle-ci est corrélée à des taux de reproduction relativement élevés, des générations à courte durée de vie, vite renouvelées, et des populations importantes. Ce sont d’autres clefs du succès des lignées qui ont franchi le passage KT. Après l’extinction des dinosaures terrestres, les tailles ont très vite augmenté, dans les premiers millions d’années du Tertiaire, et notamment chez les herbivores. La grande taille, corrélée à un métabolisme plus bas et une moindre déperdition de chaleur corporelle (et de plus grands territoires), permet en effet d’accommoder le faible rendement énergétique des végétaux (feuilles, herbes).
6. Développement du cerveau
Le cerveau des mammifères est de 4 à 10 fois plus grand que chez les reptiles, relativement à la taille du corps. Pour certains auteurs, il est aussi un élément du succès évolutif des mammifères, notamment dans le développement des stratégies adaptatives, et dans l’adaptation à des environnements hétérogènes et complexes (arbres, forêt, montagnes); il a été important dans l’évolution des carnassiers et des primates. La contrepartie est un important coût énergétique.
(EG)
Voir l’article juste paru dans Science : http://www.sciencemag.org/content/330/6008/1216.abstract