
Apport de la taxonomie intégrative à la compréhension des mécanismes à l´origine de la biodiversité des spongiaires dans les grottes sous-marines
César Augusto RUIZ PINZON
Institut Méditerranéen de Biodiversité et d’Ecologie marine et continentale
Station marine d´Endoume
Rue de la batterie des lions
13007 Marseille
cesar.ruiz (at) imbe.fr
Résumé
Dans l´objectif de comprendre les mécanismes à l´origine de la biodiversité, et l´évolution des organismes marins cavernicoles, nous avons effectué une mission de terrain d´une semaine à Santa Marta, Colombie. Cette mission d´échantillonnage représentait un site clé dans notre stratégie d’échantillonnage de la biodiversité des éponges Homoscleromorpha à l´échelle mondiale, nos sites les plus proches dans cette partie du monde se situant aux Antilles françaises et au Brésil. Différents sites ont été explorés dans la baie de Santa Marta. Une description des conditions environnementales rencontrées a été faite, et un total de 19 individus a été récolté après l’exploration d’habitats obscurs du littoral. Seulement 4 espèces d´Homoscleromorpha ont été récoltées, ce qui a tout de même permis d’ajouter une nouvelle signalisation en Colombie pour l’une d’entre elle, Plakortis angulospiculatus, et d’en signaliser une autre pour la toute première fois, Oscarella nathaliae. Cette dernière est même la toute première représentante de la famille Oscarellidae trouvée dans les eaux colombiennes. Parmi les 3 Plakinidae trouvées, Plakinastrella onkodes et Plakinastrella sp. sont deux éponges très différentes sur le plan morphologique. La première est de forme massive et de couleur jaune–crème, alors que la seconde est de forme encroûtante et de couleur grise. Au stade où en sont nos analyses de leur squelette qui est le caractère fondamental de la taxonomie des éponges, ces deux espèces partagent les mêmes types et taille de spicules. Pour conduire notre approche intégrative, la majorité des échantillons ont été fixés pour des analyses moléculaires, cytologiques et métabolomiques qui seront réalisées au cours des prochaines semaines.
Justification
La diversité du monde naturel, en particulier la profusion des espèces, a toujours été un sujet de curiosité, d´admiration et de réflexion pour les êtres humains. Aujourd´hui, il y a une conscience globale de la valeur intrinsèque de la diversité biologique et ses éléments constitutifs sur les plans environnemental, génétique, social, économique, scientifique, éducatif, culturel, récréatif et esthétique.
L’océan recouvre 71% de notre planète et représente un grand réservoir de biodiversité. Il est indispensable à plusieurs titres pour le maintien de la vie sur la terre. La plus grand partie des océans englobe une multitude d´écosystèmes qui sont encore sous explorés : zone bathypélagique, sédiments profonds, grande large, grottes sous-marines, etc.

Figure 1. Rodadero. A. Poison lion sur un corail laminaire (Agaricia sp), une espèce invasive qui s´est répandue très vite dans le Caraïbe. B. Sédiments très fins recouvrent une bonne partie du substrat et de cette Demosponge. C. les fonds rocheux offrent de nombreuses cavités et zones obscures idéales pour la recherche des éponges Homoscleromorpha. C. Oscarella nathaliae a été trouvée dans une cavité obscure.
Les grottes sous-marines sont un réservoir de biodiversité méconnu (Figure 1), qui suscitent généralement un grand intérêt grâce à leurs caractéristiques géologiques et biologiques. D’une manière générale, les peuplements des grottes sous-marines présentent un grand intérêt écologique, en particulier à cause de l’intensité des gradients physiques (lumière, circulation hydrologique, sédimentation) qui s’y déploient et du fait de la fragmentation naturelle de cet habitat. La distribution des organismes, régie par les gradients environnementaux, se traduit par une succession d’assemblages qui diffèrent de manière très marquée en fonction de la distance à l’entrée. Alors que les parties semi-obscures abritent généralement une faune très comparable à celles des substrats rocheux avoisinant, les parties obscures sont les parties qui recèlent les singularités zoologiques les plus excitantes. L’essentiel des connaissances sur les grottes sous-marines repose sur les études approfondies conduites en Méditerranée nord-occidentale, mais il reste beaucoup à apprendre de ces écosystèmes le long de la rive Sud. Par ailleurs, les explorations des grottes sous-marines démarrent à peine dans les autres régions océaniques (Pacifique, Océan Indien et Atlantique tropical).
Les éponges, Phylum Porifera, sont les métazoaires les plus anciens largement distribués sur la planète. Ce sont des organismes exclusivement aquatiques, au plan d’organisation très simple qui occupent une position basale dans l’arbre des Métazoaires, ce qui en fait des modèles de choix pour de nombreuses recherches en biologie et écologie évolutive.
Ces animaux ont colonisé tous les milieux aquatiques, dominant les paysages sous-marins dans certaines régions ainsi que certains environnements parfois jugés extrêmes. Pour la plupart filtreurs actifs puissants, les éponges sont capables de retenir un large panel de sources de nourriture, depuis la matière organique dissoute jusqu’à de petites proies eucaryotes nanoplanctoniques. Dans les grottes sous-marines ou des grands fonds océaniques, où l’absence de lumière et d’hydrodynamisme conditionnent des ressources trophiques relativement rares, les éponges ont su adapter leur stratégie de nutrition pour maximiser les prises d’énergie, développant même la carnivorie.
On s’intéresse aux éponges comme modèle d´étude, en particulier, les Homoscleromorpha, qui offrent un intérêt particulier par rapport à d’autres éponges. Tout d’abord, il s’agit d’un groupe très bien représenté dans les grottes sous-marines. Une autre caractéristique remarquable des Homoscleromorpha est sa Systématique. Chez les éponges, la composition et la structure du squelette sont les caractères taxonomiques fondamentaux. Dans le cas des éponges Homoscleromorpha, la fréquente absence de squelette a stimulé la recherche d’autres caractères, parmi lesquels des marqueurs chimio-taxonomiques. Cet approche de la taxonomie, qui vise à utiliser plusieurs types de donnes de façon complémentaire est connue comme la Taxonomie intégrative.
La Taxonomie est consacrée à la découverte de la diversité biologique et à son classement, une bonne connaissance taxonomique, voire même de la Systématique d’un groupe (relations de parentés entre organismes d’un même taxon), est un prérequis nécessaire pour de nombreuses disciplines en biologie, écologie et même en chimie des produits naturels. La Taxonomie Intégrative est aujourd’hui considérée comme l’approche la plus rigoureuse en Systématique, recommandant l’utilisation d’au minimum trois types de caractères taxonomiques indépendants. En particulier pour les éponges Homoscleromorpha, la combinaison de données de morphologie, de cytologie, d’embryologie, de génétique et de métabolomique a permis (i) démonter le « mythe de l’espèce cosmopolite Oscarella lobularis », (ii) décrire de nombreuses nouvelles espèces et résoudre des complexes d’espèces, (iii) d’élucider les relations phylogénétiques au sein de ce groupe et préciser sa position par rapport aux autres groupes d’éponges.
Ayant déjà échantillonné les Antilles Françaises, la mission de terrain en Colombie était de grande importance car elle représentait un endroit clé de la Caraïbe à proximité du continent américain. Cette mission offrait aussi l´opportunité d´améliorer la connaissance de la biodiversité des Homoscleromorpha en Colombie et d’accroître notre échantillonnage d’espèce pour améliorer notre vision de la phylogénie de ce groupe.
Méthodes
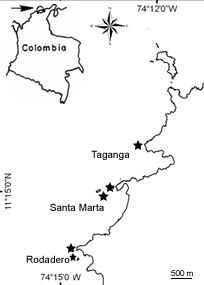
Figure 2. Localisation des sites d’échantillonnage dans la Baie de Santa Marta, au nord de la Colombie.
Site d´étude – La mission de terrain a eu lieu à Santa Marta, au nord de la Colombie, sur la côte Caraïbe. Une exploration minutieuse des cavités semi obscures et grottes a été faite de la baie de Rodadero jusqu´à Taganga (Figure 2).
Echantillonnage, traitement et conservation des éponges – Les spécimens ont été photographiés en conditions naturelles et collectés en plongée avec l´aide d´un couteau. La couleur et profondeur de chaque individu a été notée sur le terrain. Une identification préliminaire a été faite au laboratoire selon l´analyse du squelette lorsqu’il était présent. Pour chaque individu prélevé, un petit morceau a été fixé dans de l’éthanol 90% pour les analyses morphologiques et moléculaires. Pour les éponges sans squelette, un deuxième morceau a été fixé dans la glutaraldéhyde 2.5% (mélange avec eau de mer, 0.4 M de tampon phosphate et glutaraldéhyde 10% – 5 vol: 4 vol: 1 vol) pour les analyses cytologiques. Le reste de chaque individu a été lyophilisé pour l´analyse des métabolites secondaires.
Résultats et Discussion
En total, 8 plongées ont été réalisés en suivant le littoral et les fonds rocheuses jusqu´à une profondeur maximale de 30 m. En général, le substrat des sites explorées sont principalement colonisées par des Macrophytes, Coraux, Eponges. Aux endroits les plus sombres, les éponges dominent en abondance et biomasse. Différents conditions de visibilité et sédimentation ont été expérimentés le long des sites explorées. La température de l´eau était d’environ 22 °C.
Rodadero – Deux endroits ont été explorés au large du Rodadero, deux sites présentant des conditions très contrastées. Le premier site « el Morrito » est une presque île à environ 200 m du continent. Cette formation rocheuse se poursuit sous la surface par une pente douce jusqu`à 20 m de profondeur. On trouve d’énormes formations coralligènes lors des premières 10 m, les reliques d´un récif assez consolidé, car aujourd´hui ces grands madréporaires sont en grande partie morts. Les squelettes des coraux ainsi que des grandes colonies vivantes des coraux lamellaires (Fig 1A) forment un habitat composé de plusieurs cavités. La visibilité est très faible à cet endroit, principalement à cause des sédiments fins et vaseux qu´on difficulté l´exploration sur ce site (Fig 1B).
Le deuxième site exploré au Rodadero était sur la pointe nord du littoral. Le fond est rocheux, sableux par endroits, et il est dominé par les éponges. Ici, les courants sont très forts et la visibilité bien meilleure qu’à el Morrito (Fig 1C). L’habitat ne comporte que des petites cavités et fissures dans les roches, où néanmoins on a trouvé une Homoscleromorpha très rare et difficile car le plus souvent à l´abri de la lumière, Oscarella nathaliae (Fig 1D).

Figure 3. Sites explorées à Santa Marta. A. Forêt de gorgones sur le côté est d’El Morro de Santa Marta, un des sites de plongée caractérisé pour les forts courants. B. la compétition pour l´espace est très forte, même les espaces cryptiques sont fortement colonisés. C. Partie d´une épave explorée. D. échantillon 141209 SM1CR01 Plakinastrella onkodes.
Santa Marta – Deux sites ont été explorés à Santa Marta. Le premier est situé à « El Morro », un îlot à 500 m de la côte. Sous l’eau, on trouve une pente abrupte, et de forts courants (Fig 3A). On trouve des gorgones jusqu´à une dizaine de mètres de profondeur. A partir de 12 m, les madréporaires et les éponges sont les animaux qui prédominent, mais chaque espace disponible offre une grande variété de niches qui abritent des nombreuses espèces (Fig 3B).
Sur le continent, à l´extrême nord du Santa Marta, on a exploré un deuxième site, une épave qui offre un substrat artificiel (totalement recouvert) vers 17 m de profondeur (Fig 3C). Là, on a trouvé plusieurs espèces d’intérêt dans des anfractuosités, dont deux éponges Homoscleromorpha qui ont été collectées dans la majorité des sites de plongée : Plakortis angulospiculatus (Carter, 1879) et Plakinastrella onkodes Uliczka, 1929 (Fig 3D).

Figure 4. Taganga. A et B. Cette paroi verticale abrite plusieurs espèces cryptiques entre autres grâce aux conditions de faible éclairement qui y règnent. C. un deuxième site dans la même baie de Tangaga offre des conditions de lumière et de courant très différentes. D. échantillon 141210 SM2 SZ01 Plakortis angulospiculatus sur une roche exposée.
Taganga – Taganga est une baie au nord de Santa Marta. On y a exploré deux sites offrant des conditions environnementales assez différentes. Le premier est une paroi verticale qui descend jusqu´à 25 m de profondeur. On trouve tout le long de la paroi plusieurs cavités parfois totalement à l´ombre. A cet endroit, plusieurs spécimens Homoscleromorpha ont été récoltés. Certaines formes d’invertébrés massifs (éponges, madréporaires) ou dressés (octocoraux) se trouvent fixés à la paroi. Dans les petites cavités, les formes encroutâtes (principalement éponges) règnent (Fig 4A et B).
Dans le même site, mais sur un plateau situé entre 15 et 30 m de profondeur, on a exploré un site avec des formations coralligènes, substrats durs divers composés de squelettes de corail, ce qui donne un relief assez irrégulier sur le fond (Fig 4C). Dans ces endroits à l´abri de la lumière et les courants on a trouvé des éponges Homoscleromorpha (Fig 4D).
En total 19 spécimens ont été collectés dans les trois secteurs explorés entre 14 et 20 m de profondeur, généralement à l’abri d’une exposition directe à la lumière, à l´intérieur de cavités ou fissures semi-obscures.
Plakortis angulospiculatus (Carter, 1879) est une éponge massive, qui peut pendre sous la roche. Les spécimens trouvés ont une couleur marron, mesurant 5 – 10 cm2, et d’une surface légèrement rugueuse. Après la fixation dans l’alcool, aucun exsudat n’a été observé. Tous les spécimens collectés (7) présentent la même densité et les mêmes tailles de spicules : diods (70 – 110 µm long) et triods (30 – 50 µm pour la longueur des actines). P. angulospiculatus a été identifié par Zea (1987) à Santa Marta ainsi qu’à Cartagena, San Andres (Caraibe colombienne) et aux îles Bahamas, avec des variations de taille selon la localité. Une des caractéristiques diagnostiques de cette espèce est la longueur des diods toujours inférieure à 150 µm. Ainsi, elle diffère de Plakortis halichondriodes (Wilson, 1902) par la longueur des diods qui est de 100 – 190 µm, ainsi que par l´absence des triods, et une exsudation noire au contact avec l´alcool.
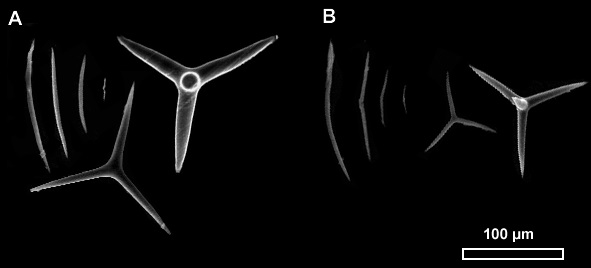
Figure 5. Image Microscope Electronique à Balayage (MEB) des spicules de deux spécimens du genre Plakinastrella. Types de spicules (gauche à droite) : diods, triods, calthropes. A. 141212 SM6CR04. B. 141209 SM1CR02.
Les spécimens identifies comme Plakinastrella onkodes correspondent à la description faite par Zea (1987) dans son étude des éponges de la Caraïbe colombienne. On a trouvé un deuxième individu qui appartient au genre Plakinastrella, mais qui présente une forme encroûtante et une couleur grise. La comparaison de la taille et des types de spicules entre tous les individus identifiés comme Plakinastrella ne montre pas de différences remarquables (longueur des diods 30-220 µm, triods 60 – 90 µm et calthropes 50 – 190 µm) (Fig 5). Une espèce de Plakinastrella en cours de description, collecté en Martinique a une morphologie externe très semblable au spécimen gris encroûtant, néanmoins la taille des spicules est légèrement inférieure (diods 40 – 140 µm ; triods 20 – 60 µm), et elle ne possède pas des clathropes.
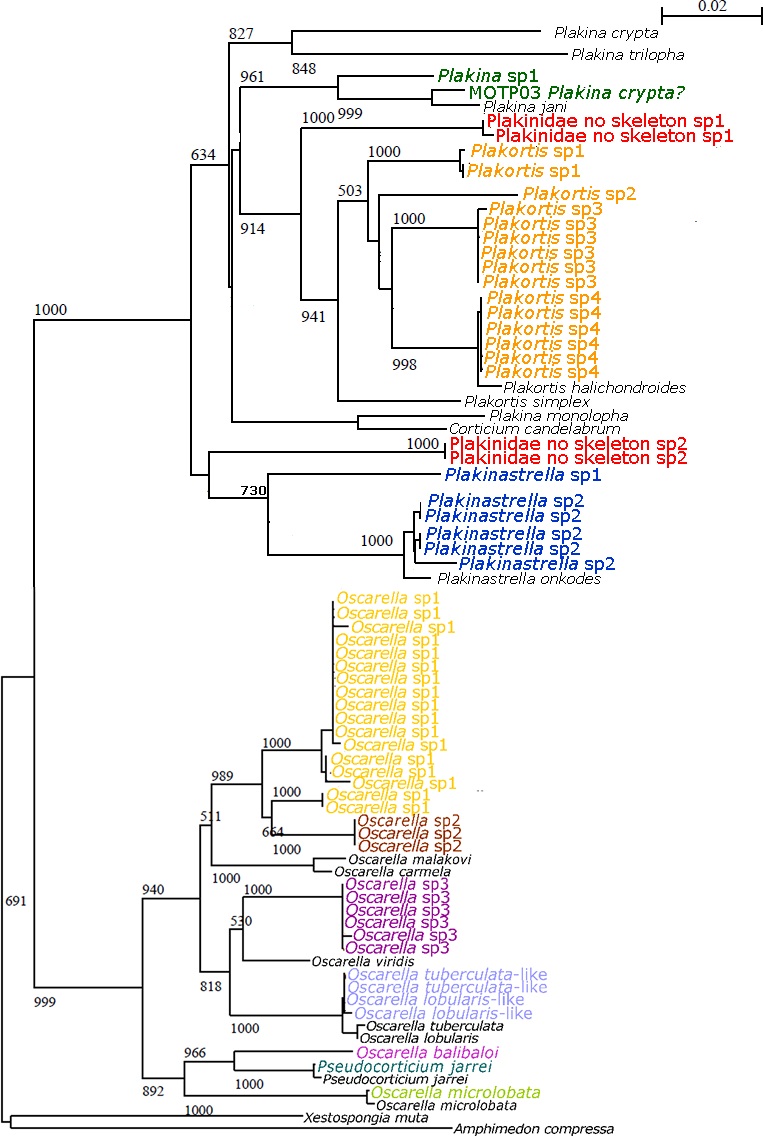
Figure 6. Arbre phylogénétique selon l´analyse de gène CO1 des Homoscleromorpha collectées lors de la thèse en cours (individus en couleur). En noir, séquences téléchargées depuis Genbank.
La taille et densité des spicules étant des caractères variables chez les Homoscleromorpha, l´analyse chimique pourrait apporter des caractères importants pour la détermination et limitation des espèces. Dans la présente étude, plusieurs spécimens ont été lyophilisés pour des postérieures analyses chimiques, qui, couplées aux données moléculaires, aideront à mieux comprendre les relations phylogénétiques entre les Homoscleromorpha (Fig 6).
Apparemment la présence / absence du squelette n´est pas un caractère diagnostique pour définir les deux familles au sein des Homoscleromorpha. Par exemple, Oscarella nathaliae Ereskovsky, Lavrov & Willens, 2014, est une espèce qui par l´absence de spicules et l´étude cytologique a été classifie au sein de la famille Oscarellidae (éponges sans spicules). Nous sommes en train d´analyser 6 spécimens de O. nathaliae collectés en La Martinique et La Guadeloupe, parmi ses individus, nous avons réussi à amplifier (CO1) et séquencer un seul spécimen dont leur séquence génétique se situe au sein de Plakinidae (éponges avec spicules), la deuxième famille des Homoscleromorpha. Grace aux deux spécimens collectés lors de cette mission, de nouvelles séquences peuvent être espérées, ce qui permettra de préciser la position de cette espèce dans la classification des Homoscleromorpha.
Remerciements
Je tiens à remercier Sven Zea pour son aide et accompagnement pendant cette mission de terrain, et l’Instituto de Investigaciones Marinas y Costeras Invemar pour le soutien logistique apporté. Merci également à la Société Française d´Ecologie pour la bourse de terrain attribuée en 2014. Cette mission a également été partiellement financée par le Laboratoire International Associé LIA MARRIO (Plongées, vol intérieur) et par le programme ECOS-Nord C12S02 (Vol international).
Références
CARDENAS P., PEREZ T., BOURY-ESNAULT N. 2012. Sponge systematics facing new challenges. Advances in Marine Biology 61: 79-209.
DAYRAT B., 2005. Towards integrative taxonomy. Biological Journal of the Linnean Society 85, 407–415.
ERESKOVSKY A.V., IVANISEVIC J., PEREZ T. (2009). Overview on the homoscleromorpha sponges diversity in the Mediterranean. Proceedings of the 1st Symposium on the Coralligenous and other calcareous bio-concretions of the Mediterranean Sea, pp. 89-95.
GAZAVE E., LAPÉBIE P., ERESKOVSKY A.V., VACELET J., RENARD E., CÁRDENAS P., BORCHIELLINI C. 2012. No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera. Hydrobiologia 687 : 3-10.
HARMELIN .G., VACELET J.,VASSEUR P., 1985. Les grottes sous-marines obscures : un milieu extrême et un remarquable biotope refuge. Téthys 11, 214-229.
IVANISEVIC J., THOMAS O.P., LEJEUSNE C., CHEVALDONNE P., PEREZ T. 2011. Metabolic fingerprinting as an indicator of biodiversity: towards understanding inter-specific relationships among Homoscleromorpha sponges. Metabolomics 7: 289–304.
MURICY G., BOURY-ESNAULT N., BÉZAC C., VACELET J. 1996a. Cytological evidence for cryptic speciation in Mediterranean Oscarella species (Porifera, Homoscleromorpha). Canadian Journal of Zoology 74, 881–896.
MURICY, G., SOLÉ-CAVA, A. M., THORPE J. P., BOURY-ESNAULT N. 1996b. Genetic evidence for extensive cryptic speciation in the subtidal sponge Plakina trilopha (Porifera: Demospongiae: Homoscleromorpha) from the Western Mediterranean. Marine Ecology Progress Series 138, 181–187.
PEREZ T., IVANISEVIC J., DUBOIS M., PEDEL L., THOMAS O. P., TOKINA D., ERESKOVSKY A.V. 2011. Oscarella balibaloi, a new sponge species (Homoscleromorpha: Plakinidae) from the Western Mediterranean Sea: cytological description, reproductive cycle and ecology. Marine Ecology 32(2): 174–187.
REVEILLAUD J., ALLEWAERT C., PEREZ T., VACELET J., BANAIGS B., VANREUSEL A. 2012. Relevance of an Integrative Approach for Taxonomic Revision in Sponge Taxa: Case Study of the Shallow-water Atlanto-Mediterranean Hexadella (Porifera, Ianthellidae, Verongida). Invertebrate Systematics 26 (3): 230-248.
RUIZ C. 2012. Taxonomie intégrative des éponges Homoscleromorpha : approches cytologique, chimique et moléculaire. Master 2, Aix-Marseille Université.
VACELET J., BOURY-ESNAULT N. & HARMELIN J.G., 1994. Hexactinellid Cave, a unique deep-sea habitat in the scuba zone. Deep-Sea Research I 41, 965-973.
ZEA S. 1987. Esponjas del Caribe colombiano. Cátalogo Científico, Bogotá. 226 p.
Commentaires récents