Compétition acoustique et évolution du chant en contexte insulaire.
Aloïs Robert
Centre d’Écologie Fonctionnelle et Évolutive ,
1919, route de Mende, Montpellier.
Email: alois.robert@cefe.cnrs.fr
Co-auteurs : Aloïs Robert(1), Sacha Julien(1), Doris Gomez(1), Thierry Lengagne(2), Martim Melo(3), Rita Covas(3) & Claire Doutrelant(1).
(1) CEFE, UMR 5175, Montpellier
(2) LEHNA, UMR 5023, Lyon
(3) CIBIO, Université de Porto, Portugal
Contexte et enjeux
La communication étant cruciale pour la survie et la reproduction des individus, connaître et comprendre les processus qui régulent la communication des êtres vivants est un enjeu important en écologie. Dans cette optique, l’écologie des communautés acoustiques représente un angle d’étude prometteur en permettant d’évaluer l’importance de ces processus à large échelle [2]. De nombreuses études ayant classiquement porté sur l’influence de la végétation sur le chant [1, 9], la structuration globale des signaux acoustiques par l’environnement est bien comprise. Cependant, les mécanismes conduisant à la structuration fine de ces signaux restent largement méconnus [2]. Ce projet propose d’étudier une source potentielle de cette structuration fine: la compétition sensorielle entre espèces sympatriques. Cette force est la résultante directe de la complexité des communautés animales.
Les espèces sympatriques qui communiquent par signaux sonores doivent se partager l’espace acoustique défini par l’environnement dans lequel les sons se propagent [10], environnement qui par ses caractéristiques physiques définit un nombre restreint de signaux optimaux pour communiquer. L’espace acoustique peut être ainsi considéré comme une ressource limitée pour laquelle les espèces rentrent en compétition [11]. Cette compétition acoustique a été montrée comme étant une force évolutive majeure pour les signaux acoustiques et la diversification des espèces [12, 13], mais son importance pour la structuration des communautés acoustiques reste très peu étudiée. Au niveau écologique, il a été démontré qu’à habitat identique, la compétition interspécifique et la diversité spécifique plus faibles sur les îles que sur les continents amenaient à un élargissement des niches écologiques [14, 15]. Notre hypothèse est que le même processus intervient sur la niche acoustique des espèces, une hypothèse récemment suggérée par des analyses que nous avons faites sur l’évolution des chants d’oiseaux dans les îles [16].
Nous avons ici mesuré la compétition pour cette espace sur le terrain en mesurant le chevauchement temporel des espèces co-chantant et la niche acoustique de chaque espèce. La prédiction est que la compétition, plus sévère dans les communautés continentales et plus riche en espèces, conduit à une congestion de niche, c’est à dire à une réduction de l’espace acoustique utilisé par chaque espèce.
Méthodes
Nous avons mesuré le chevauchement temporel des chants d’espèces insulaires et l’avons comparé au chevauchement temporel des chants de leurs plus proches espèces continentales. Le chevauchement temporel est notre proxy pour la compétition acoustique. Nous avons mesuré la gamme de fréquence des chants du set d’espèces insulaires et l’avons comparé au chevauchement temporel des chants de leurs plus proches espèces continentales. Cet gamme de fréquence est un indicateur de la taille de la niche acoustique des espèces. Afin de nous assurer que le filtrage par l’environnement est le même dans les sites comparés nous avons procédé à des expériences de propagation.
Deux paires île/continent ont été étudiées : São-Tomé/Cameroun et Madère/Sud de la France. Nous ne parlerons ici que de l’étude de la paire Madère/Sud de la France dont la mission de terrain a été effectuée grâce au financement de la SFE.
En Mars 2017 une mission a été effectuée à Madère (Aloïs Robert, Claire Doutrelant et un étudiant de M1 : Sacha Julien). En avril 2017, une dernière mission a été effectuée près de Montpellier (Aloïs Robert, Malicia Besnard et Sacha Julien) pour avoir un point continental en Europe.

Illustration 1 : Un chemin à travers la Laurisylve, forêt primaire (datant du Miocène) de l’île de Madère.
1. Enregistrement de l’espace sonore utilisé
Le paysage sonore (i.e. espace acoustique utilisé) est multidimensionnel (paysage de fréquence, de durée, de syntaxe avec leurs composantes temporelles). Pour le caractériser, nous avons utilisé des boîtes d’enregistrement automatique SM3. Les enregistrements par ces boîtes permettait en outre de quantifier le nombre d’espèces en compétition pour l’espace acoustique dans chaque communauté. Le nombre d’espèces présentes et le nombre d’espèces co-chantant au même moment du matin sont relevées à partir de ces enregistrements. Chaque communauté a été échantillonnée dans trois répliquâts pour lesquels un total d’une quinzaine d’enregistrements omnidirectionnels espacés de plus de 200m étaient réalisés avec les boîtes SM3.

Illustration 2 : Aloïs portant une boîte SM3 à travers la laurisyle à Madère.
Sur les boîtes SM3, les enregistrements commencent dans le chorus matinal et durent 3h30. Pendant cette période seront identifiées toutes les 15 min les espèces co-chantant sur une période de 3 min. Ainsi 42 min de chant seront analysées par matinée d’enregistrement et par boîte SM3, chaque habitat étant caractérisé par au moins 8 boîtes.
2. Enregistrement des chants produits par les espèces focales avec des micros unidirectionnels
Ce travail nous permet de mesurer la niche acoustique des espèces focales. Nous avons d’abord échantillonné la forêt primaire de l’île (laurisylve voir illustrations 1) puis avons échantillonné de vieilles forêts de chênes verts sur le continent (ces deux types de forêts présentent des particularités physico-acoustiques communes ; voir partie 3). Au moins 20-30 individus de chaque espèce focale ont été enregistrés sur chaque site. Ce chiffre a dû être réduit pour l’analyse acoustique car de nombreux enregistrements étaient bruités. Par ailleurs pour qu’un enregistrement puisse être utilisé il faut que l’enregistrement présente un bon rapport signal/bruit et que les sons ne présentent aucun recouvrement avec d’autres espèces d’oiseaux. Les espèces étudiées, le nombre d’enregistrements et le nombre d’oiseaux qui ont pu être analysés au final sont présentés dans le tableau 1.
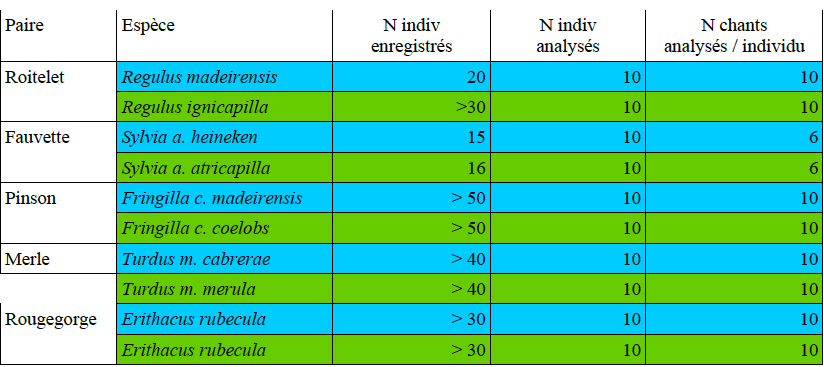
Tableau 1 : Echantillonnage réalisé pour les espèces focales, en bleu les espèces de l’île, en vert les espèces continentales.
Sur ces chants, nous avons pris les mesures suivantes (Fig. 1) pour caractériser la niche acoustique de l’espèce : Mesures syntaxiques : nombre total de notes par chant, nombre de notes différentes par chant, complexité du signal (richesse = nombre de notes différentes * versatilité = nombre de transitions entre les différentes notes); Mesures temporelles : durée du chant (sec), nombre de notes par seconde ; Mesures fréquentielles : fréquence minimum, maximum, bande de fréquence, pic de fréquence, quartile à 25% et 75%.
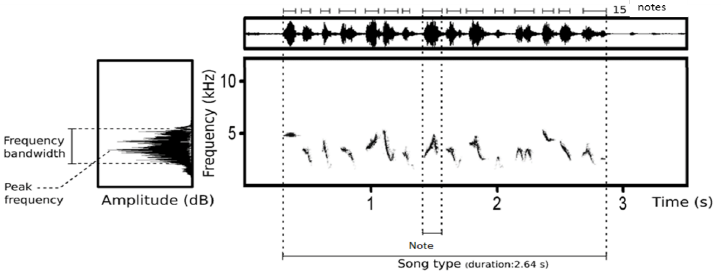
Figure 1.
3. Réalisation des expériences de propagation du son
Pour minimiser l’effet de l’habitat végétal sur les caractéristiques du son des espèces focales nous avons travaillé dans les mêmes habitats. Cependant il fallait vérifier ce présupposé. Pour ce faire nous avons suivi le protocole de Thierry Lengagne (LEHNA, Lyon). La propagation nous permettra de vérifier la similarité des contraintes acoustiques des habitats comparés et de déterminer si l’espace acoustique disponible est le même au sein de chaque habitat comparé (l’espace dans lequel les sons se transmettent bien à 30 m dans chaque habitat). Pour ces expériences, Thierry Lengagne a fait construire un haut-parleur permettant de faire des repasses de qualité. Ce haut parleur, un Monacor SPH60X associé à un amplificateur Dynavox DA-30 et un lecteur WAV, était placé à 2,4m du sol et repassait ainsi des sons à 2,5m du sol à un niveau de 80db à 1m. Deux types de son étaient repassés sur le terrain: un bruit blanc contenant toutes les fréquences et une modulation de fréquence ascendante qui balaye la gamme 200Hz-20000Hz. Ces sons étaient ré-enregistrés à 1m, 10m, 20m et 30m. Au labo, le signal est analysé par zero-crossing afin de suivre l’évolution de la fréquence dans le temps. Au fur et à mesure que le signal est altéré par la dissipation sphérique de l’énergie et par les obstacles rencontrés lors de la propagation (phénomènes de réflexion et de réfraction sur les obstacles) il n’est plus possible de suivre le signal. Il est alors possible de calculer un taux de ressemblance du signal en mesurant le pourcentage du signal qui n’est plus suivi par le zero-crossing.

Illustration 3 : Sacha en train d’effectuer les dernier réglages avant les mesures de propagation.
Résultats préliminaires
1. Chevauchement temporel et compétition acoustique
Pour l’instant, nous avons utilisé le chevauchement temporel du chant comme proxy de compétition acoustique. Nous trouvons que le nombre moyen d’espèces sympatriques avec qui les espèces insulaires présentent un chevauchement temporel est moins important que celui des espèces continentales. De plus ces résultats sont retrouvés à l’échelle de la communauté comme à l’échelle de nos espèces focales (Fig. 2).
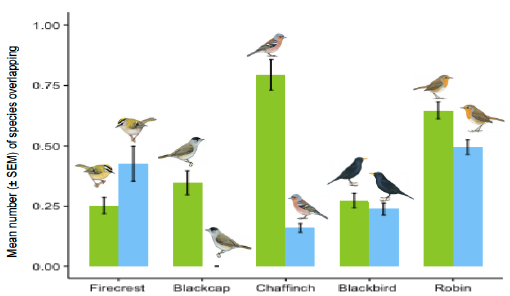
Figure 2 : Nombre moyen d’espèces chevauchant temporellement chaque chant des espèces focales (continentales en vert, insulaires en bleu).
Ce proxy de compétition acoustique est cependant insuffisant et nous le couplerons prochainement avec une mesure du chevauchement spectral. En effet, deux espèces dont les chants se chevauchent temporellement mais pas en fréquence ne sont pas réellement susceptibles d’entrer en compétition de même que deux espèces dont les chants se chevauchent spectralement mais qui s’évitent temporellement.
2. Gammes de fréquences des espèces focales
Concernant les variations de la gamme de fréquence au sein des différentes paires d’espèces focales, les résultats ne montrent pas de tendances claires (Fig. 3). Pour deux espèces sur cinq on a une augmentation significative de la gamme de fréquence chez l’espèce insulaire alors que pour une espèce sur cinq on a une diminution. En modélisant la variation de la gamme de fréquence en fonction de la population (continent/île) par un modèle mixte incluant la structure phylogénétique on ne trouve pas d’effet significatif du type de population sur la gamme de fréquence.
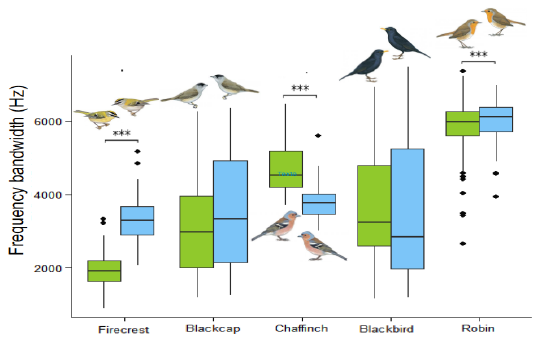
Figure 3 : Gamme de fréquences des espèces focales (continentales en vert, insulaires en bleu).
3. Propagation dans les différents milieux échantillonnés
Les expériences de propagation montrent des profils extrêmement similaires pour les milieux échantillonnés suggérant que les contraintes environnementales sur les gammes de fréquences utilisés sont très proches.
Conclusion
Ainsi, malgré une diminution de la compétition acoustique dans l’île, les espèces insulaires ne présentent pas de diminution de leur gamme de fréquence depuis leur colonisation. Ce travail prend place au sein d’une étude à plus grande échelle incluant une comparaison de la compétition acoustique et de gammes de fréquences d’une paire îles/continent en milieu équatorial (Sao-Tomé/Cameroun). Ce travail a montré que en milieu équatorial la compétition pour l’espace acoustique i) est plus grande que celle retrouvé dans la paire tempéré (Madère/France) ii) où elle est plus grande sur le continent que sur l’île. Il a aussi montré que les gammes de fréquences des espèces insulaires étaient significativement plus larges que celles des chants des espèces continentales suggérant un relâchement de caractères. Nous pensons que ce relâchement fait suite à la colonisation de l’île par des espèces continentales car elles endurent alors une moindre compétition acoustique du fait du nombre d’espèces sympatriques restreint. Comme la compétition acoustique en milieu tempéré est moindre que celle retrouvé en milieu équatorial on peut expliquer les résultats que l’on trouve ici par le fait que, sous des latitudes tempérés, même en milieu continental, il n’y a pas de congestion de niche acoustique. Cela sera étudié en utilisant des proxy de compétition acoustique plus précis.
De manière plus large, ce travail de terrain a mis en évidence le manque de données de base relatives à la phénologie des espèces sur l’île de Madère et en Macaronésie en général. En effet, il nous a été très difficile de cibler avec certitude les dates de reproduction des espèces insulaires pour être certains que ces espèces seraient chanteuses au moment de la mission de terrain. À ma connaissance il n’existe pas d’ouvrage de référence sur la biologie de ces espèces ou sous-espèces insulaires qui sont pourtant assez différentes de leurs apparentés continentaux. Cela montre encore une fois que l’écologie de terrain est loin d’être obsolète ainsi que toute l’importance de bourses de terrain telles que celle de la Société Française d’Écologie.
Références
[1] Smith et al., 2013 Evol Appl. 6, 865-874. [2] Pijanowski, et al, 2011, BioScience, 6, 203-216. [3] Tobias et al., 2014 PNAS. 111, 1020–1025. [4] Weir et al., 2011 Proc. Biol. Sc. 278, 1713-1720. [5] Le Saout et al, 2013Science, 342, 803-805 [6] Lengagne, T., et al. (2015). Plosone. [7] Doutrelant, C., et al. In press. Ecology Letters. [8] Melo, et al. (2011). Mol. Ecol. 20, 4953-4967. [9]Wiley, 1991Am. Nat. 138, 973-993 . [10]Luther 2009 Behav Ecol, 20:864–871 [11] Hart et al., 2015 Behav Ecol, 20:1-4. [12] Grether et al. 2009, Bio. Rev., 84, 617-635; [13] Grant & Grant. 2010 PNAS.107, 20156–20163. [14] MacArthur, et al. 1972 Ecology 53:330–342 . [15] Blondel et al., 1988 Ecology 69:1899–1917. [16] Morinay, et al. 2013Ecol. Evol ; 3: 5127–5140 . [17] Lengagne, T. & Slater, P. J. (2002). Proc.Roy. Soc. B, 269, 2121-2125
Commentaires récents