La Société Française d’Ecologie (SFE) vous propose le regard de de Alexandre Robert sur les petites populations.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
Petites populations et vortex d’extinction
Alexandre Robert
Chercheur au Muséum National d’Histoire Naturelle, CERSP, UMR 7204
( Fichier PDF )
Regard R9, édité par Anne Teyssèdre
———
Mots clés : Extinctions, populations, mécanismes, génétique, dynamique, hasard, mutations, vortex d’extinction.
———
L’étude des causes qui mènent les espèces vivantes à la rareté ou à l’extinction sont au centre des sciences de la conservation. Une des plus célèbres classifications des causes de raréfaction des espèces est celle proposée par le biologiste J. Diamond, qui définit quatre causes majeures de déclin des espèces : la modification des habitats, la surexploitation, l’introduction d’espèces exogènes dans les écosystèmes, et enfin la perturbation des réseaux écologiques (où par exemple, la disparition d’une proie va affecter la survie de son prédateur) (Diamond 1989). Cette classification déjà ancienne reste d’actualité dans la mesure où elle regroupe l’essentiel des causes de déclin directes ou indirectes d’origine humaine, et notamment le changement climatique qui est une cause majeure de modification des habitats. Mais suffit-il de connaître les causes de déclin des espèces pour pouvoir les protéger ? Que se passe-t-il entre le déclin et l’extinction d’une espèce ?

© Alexandre Robert
L’écologie et la génétique nous enseignent que lorsqu’une population devient petite, un certain nombre de processus démographiques, génétiques, physiologiques ou comportementaux se trouvent perturbés. La constitution de groupes suffisamment grands ou denses est en effet un moyen d’assurer les interactions entre individus nécessaires à la reproduction, de rendre l’environnement physico-chimique plus favorable (en acidifiant les sols, en régulant la température…), de chasser plus efficacement ou au contraire d’échapper aux prédateurs. On observe ainsi chez de nombreuses espèces que les petites populations ne sont pas capables de croître aussi rapidement que les grandes, ou que leur croissance est extrêmement variable dans le temps. L’objet de cet article est d’expliquer et d’illustrer certains de ces phénomènes. On se concentrera sur les mécanismes démographiques et génétiques qui sont les plus universaux, et dont les effets et leurs interactions contribuent largement à ce que l’on nomme le vortex d’extinction.
L’incertitude démographique
La démographie est une discipline qui s’intéresse à l’abondance des populations et à la répartition des individus selon des classes d’âge, de sexe, etc. Dans un environnement donné, une population est caractérisée par ses taux de natalité, mortalité et dispersion, qui déterminent la dynamique globale et moyenne de la population (stabilité, croissance,…). Mais si l’on regarde les phénomènes démographiques à l’échelle individuelle, on s’aperçoit que tous les individus n’ont pas le même destin. Certains meurent vieux, d’autres jeunes. Certains laissent beaucoup de descendants, d’autres aucun. Ces différences sont dues au fait que les individus d’une population diffèrent les uns des autres dans leur capacité à se reproduire, survivre, et au fait que, même pour des individus ayant tous le même potentiel, le destin de chacun (mort, reproduction…) est considéré comme un événement aléatoire. Le hasard n’est pas vu ici comme un mécanisme biologique ayant une existence propre, mais comme la résultante de l’imprédictibilité de certains phénomènes pour nous, observateurs.
Prenons l’exemple fictif d’une plante hermaphrodite annuelle, pollinisée par des abeilles et dont les graines sont dispersées – et consommées ! – par des passereaux. Chaque année, quand les conditions environnementales sont favorables, chaque individu est fécondé et produit une centaine de graines dont en moyenne cinq germent et 1,3 survivent jusqu’à l’âge adulte, avant de se reproduire à leur tour et mourir. Dans le cadre d’une grande population, ces taux moyens de germination et de survie sont suffisants pour assurer la croissance de la population, qui peut d’ailleurs être facilement estimée (de la même façon que nous pouvons approximer le résultat d’un grand nombre de lancers de pièce « pile ou face »).
En revanche, dans le cadre d’une petite population (voire quelques individus), la dynamique globale va dépendre d’un très faible nombre d’événements de germination et de mortalité ; il est alors possible que la population dévie fortement de son comportement moyen attendu. Ceci a deux conséquences majeures. La première, c’est que nous ne sommes plus capables de prédire correctement sa dynamique. Bien que l’échec à la germination et la mortalité aient des causes réelles (micro habitat défavorable, hasard des pollinisations, prédation, …), ils demeurent pour nous imprédictibles à l’échelle individuelle (tout comme l’est un seul lancer de pièce). La deuxième, c’est que la population peut s’éteindre. Même si la taille de population n’a aucune influence sur les taux démographiques, et même si la population a en moyenne la potentialité de croître, le risque d’extinction (c’est-à-dire le risque qu’aucun individu n’atteigne l’âge adulte) est plus élevé dans une petite que dans une grande population.
Les mécanismes de détérioration génétique
Les caractéristiques des espèces menacées (petites populations, souvent fragmentées), sont favorables à l’apparition de problèmes génétiques, dont les principaux sont décrits ci-dessous.

© Anne Teyssèdre
La dépression de consanguinité. La consanguinité d’une population résulte de l’accouplement d’individus qui se ressemblent génétiquement (par exemple, provenant de la même famille). Chez de nombreuses espèces, de tels croisements consanguins sont rares tant que l’effectif de la population (i.e. le nombre d’individus) est grand. Mais lorsque l’effectif diminue, la population devient génétiquement homogène et les croisements consanguins entrainent l’expression de mutations défavorables qui affectent la survie ou la fécondité des individus.
La fonte mutationnelle. Dans les grandes populations, les mutations désavantageuses qui se produisent en permanence et se transmettent sont éliminées par le processus de sélection naturelle. Dans les petites populations, ce processus de sélection n’est plus aussi efficace et aboutit à l’accumulation de mutations délétères, qui peu à peu, vont réduire la capacité de la population à croitre.
La perte de potentiel évolutif. Sur le long terme, l’environnement des espèces est fluctuant, pour des raisons astronomiques, climatiques, géologiques, biotiques et, de plus en plus souvent, anthropiques. Pour s’adapter à ces changements perpétuels, les espèces peuvent puiser dans la diversité de leur patrimoine génétique. Lorsque la taille d’une population diminue, cette diversité diminue, ainsi que la capacité à s’adapter à d’éventuels changements environnementaux.
Contrairement aux phénomènes démographiques évoqués plus haut, ces processus peuvent avoir des conséquences durables, voire irréversibles, même lorsque les populations ne sont réduites que de façon temporaire (on parle alors de goulot d’étranglement), comme dans le cas des espèces qui ont été fortement exploitées ou persécutées, avant d’être protégées légalement. Le bison américain a par exemple vu sa population fondre de plus de 60 millions à 750 individus à la fin du 19ème siècle. Depuis il est protégé et compte désormais plusieurs centaines de milliers d’individus.
On sait que lorsqu’une population isolée présente des problèmes de détérioration génétique, , l’arrivée de nouveaux individus extérieurs à la population peut avoir des effets bénéfiques spectaculaires. Mais lorsque la population menacée comporte les derniers représentants de l’espèce, la restauration de la variation génétique perdue nécessiterait que la population se maintienne à un effectif élevé pendant de nombreuses générations, ce qui est généralement impossible. Ainsi, les goulots d’étranglement sont des phénomènes qui peuvent être démographiquement réversibles, mais génétiquement irréversibles.
Un vortex d’extinction
Le terme « vortex d’extinction », que l’on doit à M. Gilpin et M. Soulé (Gilpin & Soulé 1986), fait référence aux rétroactions positives (renforcement mutuel entre différents processus) qui peuvent intervenir lorsque les populations déclinent, par exemple, du fait de l’altération de leur habitat. Ces processus peuvent avoir diverses origines – démographique, physiologique, comportementale… -, mais on va ici se focaliser sur l’interaction démographie-génétique, formalisée par le généticien M. Lynch et ses collaborateurs (Lynch et al. 1995).
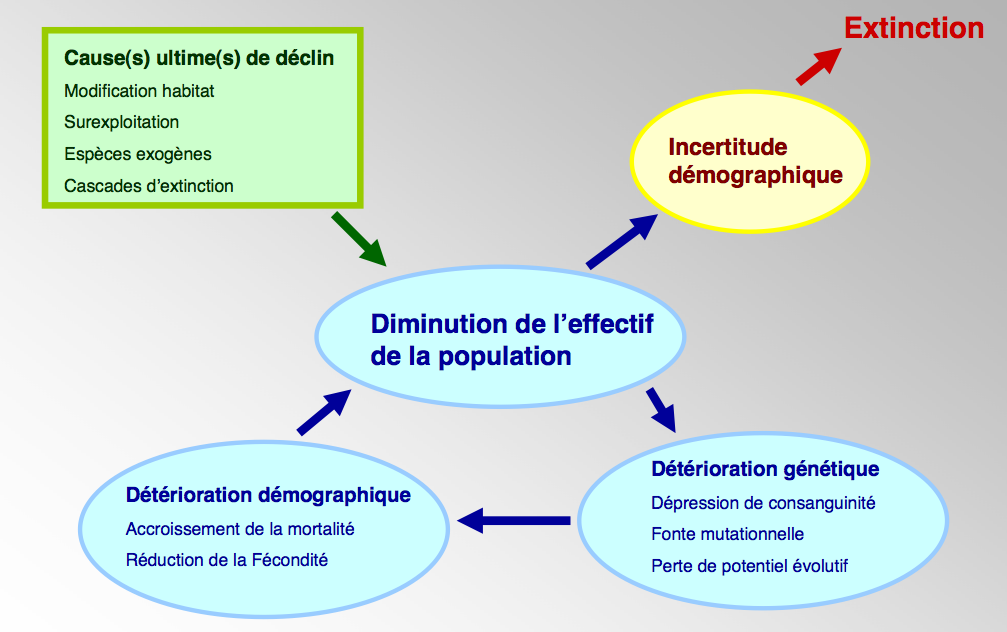
Le mécanisme est simple. Une population isolée, de petite taille, accumule progressivement des mutations désavantageuses – c’est la fonte mutationnelle. Chacune de ces mutations peut réduire par exemple la fécondité des femelles, mais sans influer sur la taille de la population, qui reste constante (il s’agit de la taille imposée, par exemple, par le nombre de sites de reproduction disponibles).
Lorsque le nombre de mutations désavantageuses devient trop grand, la fécondité moyenne devient trop faible pour compenser la mortalité naturelle, et la population se met à décliner. Nous entrons dans le vortex d’extinction, où les problèmes génétiques (mutations désavantageuses) et démographiques (taille trop faible et déclinante) s’amplifient mutuellement. A mesure que la taille diminue, les problèmes génétiques deviennent plus aigus (accumulation encore plus rapide de mutations), ce qui fait diminuer la taille de population encore plus rapidement et amplifie les problèmes liés à l’incertitude démographique (déséquilibre dans le sex-ratio, etc. voir schéma 1). Ce vortex conduit théoriquement à l’extinction de la population.
Conclusion
Le vortex d’extinction des petites populations nous enseigne que celles-ci ne sont pas simplement des modèles réduits de grandes populations. Dans une petite population, même lorsqu’aucun processus biologique n’est affecté à l’échelle individuelle (comme dans le cas de l’incertitude démographique), le risque d’extinction augmente intrinsèquement par rapport à une grande population. A ces phénomènes aléatoires s’ajoutent de nombreux mécanismes qui affectent les petites populations à l’échelle individuelle et populationnelle, tels que les problèmes génétiques et les effets Allee (Courchamp et al. 1999). L’ensemble de ces phénomènes sont susceptibles de s’amplifier mutuellement dans un vortex d’extinction.
Il est évident que si les facteurs humains associés au déclin des espèces – tels ceux qui entrent dans la classification de Diamond – ne sont pas écartés, l’étude du vortex d’extinction ne sert à rien. Mais, à l’inverse, même lorsque les causes ultimes de déclin sont écartées, les problèmes associés aux faibles effectifs sont susceptibles de persister durablement. L’écologie évolutive nous fournit des outils pour atténuer les effets délétères de ces phénomènes lorsqu’ils affectent des populations locales (par exemple en transférant des individus extérieurs vers les populations les plus menacées) mais elle nous enseigne également que ces effets sont virtuellement irréversibles lorsqu’ils affectent une espèce dans son ensemble. Le maintien des espèces à des effectifs suffisamment importants reste donc une des solutions à adopter pour éviter qu’elles n’entrent dans le vortex.
Bibliographie
Diamond J. (1989) Overview of recent extinctions. In: Conservation for the Twenty-First Century (eds D.Western and M. C.Pearl), pp. 37–41. Oxford University Press, New York.
Gilpin ME, Soulé ME (1986). « Minimum Viable Populations: Processes of Species Extinction ». In M. E. Soulé. Conservation Biology: The Science of Scarcity and Diversity. Sinauer, Sunderland, Mass. pp. 19–34.
Lynch, M., J. Conery, and R. Bürger. 1995. Mutational meltdowns in sexual populations. Evolution 49:1067–1080.
Courchamp, F., T. Clutton-Brock, and B. Grenfell. 1999. Inverse density dependence and the Allee effect. Trends in Ecology and Evolution 14:405–410.
Ce regard sur un phénomène compliqué est très bien vulgarisé et par conséquent très compréhensible. Merci Dr Robert!
Puisque l’on connait les concésquences, comment faire pour revenir en arrière et que le processus de déclins soit inverser ?
Merci pour ces explications très pédagogiques. Je ne comprends pas le sens que vous donnez à la notion de hasard. Est-ce vraiment rigoureux de parler de hasard, le hasard a-t-il un sens en sciences, et si oui lequel ? cdt, JCM
Bonjour a tous, cette idee de plate-forme de discussion entre scientifiques et citoyens est geniale. Si je peux me permettre de repondre a Coba a propos de l’utilisation du terme hasard. Le terme hasard est en general utilise pour identifier les lacunes dans l’identification des relations de cause a effet d’un evenement. Par definition meme la science est faite pour combattre le hasard, ou du moins, pour identifier les relations de causes a effet d’un evenement particulier. Par exemple, la derive genetique est l’une des quatre principales forces evolutives: il s’agit de l’evolution genetique aleatoire qui est du au tri aleatoire (du au hasard) des individus et des genes. Par exemple, parmi des millions de spermatozoides qui se lancent dans la course pour la vie, un seul (voire deux parfois) reussira(rons). Utiliser ce terme en science est a mon sens tout a fait rigoureux, lorsqu’un evenement ou un mecanisme ne peut etre pour l’instant explique.
Réponse à Coba :
ma réponse est de la même teneur que celle de Twaro (merci de vos messages) :
Oui le hasard a certainement un sens en science, voire plusieurs. Sa définition dépend à la fois des phénomènes considérés, de l’échelle, et de l’observateur. Ce sont surtout les philosophes et les physiciens qui ont écrit sur le sujet, moins les biologistes (mais voir le livre de J. Monod, le hasard et la nécessité ; et comme le souligne Twaro, l’aléa joue un rôle clé en biologie évolutive, ou tout au moins dans la représentation que l’on se fait des phénomènes évolutifs). En physique quantique, certains postulats considèrent le monde comme indéterministe : il existerait un hasard authentique et irréductible (i.e., l’absence de déterminisme n’est pas qu’apparent, il est indépendant de la mesure). Dans ce contexte, une même cause ne va pas forcément produire les mêmes effets et l’état d’une particule, par exemple, peut être déterminé par heu… rien. Mais comme je ne suis pas physicien, je risque de dire énormément de bêtises si je continue sur cette voie.
Heureusement, nous nous situons ici dans un cas plus classique en sciences, où le « hasard » est lié à l’observation. Dans le contexte de la dynamique des populations, je considère qu’à notre échelle « macro », les événements démographiques individuels qui m’intéressent sont déterministes, dans le sens où chaque événement (mort, naissance…) est déterminé par une ou plusieurs causes (autrement dit, à mon échelle, dans le monde réel, le hasard n’existe pas). C’est la description que j’en fais qui fait intervenir le hasard. L’image que j’ai de ma population est une image probabiliste, faite de statistiques imparfaites (notamment, mes paramètres démographiques, qui sont le plus souvent, des moyennes, or les moyennes n’existent pas dans la vraie vie, il n’y a que des événements). Si mes statistiques sont imparfaites, c’est parce qu’elles sont intégratives (je décris des événements individuels à l’échelle d’une population), et parce que ma vision du monde est imparfaite : je ne suis pas capable de connaître, de comprendre, de décrire (et donc de modéliser) l’ensemble des chaines de causalités qui aboutissent à tel ou tel événement, comme la mort d’un individu ou le fait que ma pièce tombe du coté pile.
En réponse à Twaro, je dirais que la science n’est pas faite pour combattre le hasard mais plutôt pour l’intégrer dans ses analyses, synthèses et prédictions [sur le fonctionnement des systèmes physiques, biologiques, écologiques, etc..].
Bonjour,
Tout d’abord, je trouve aussi cette idée de plateforme très intéressante par les échanges qu’elle peut impliquer.
Je ne suis pas vraiment d’accord sur ce qui est dit dans les réponses sur le hasard. La diversité génétique des populations évoluent sous la contrainte de différentes forces évolutives dont la sélection naturelle et la dérive génétique qui vont toutes les deux éliminer certains individus. Ces deux phénomènes ont donc la même conséquence : réduire le nombre d’individus ou le nombre de descendants dans une population.
Cependant une énorme différence existe entre ces deux mécanismes. En effet, la dérive élimine les individus au hasard sans relation avec le patrimoine génétique de ces individus alors que la sélection (par définition) va éliminer des individus du fait d’un patrimoine génétique « délétère » en partie. Le fait qu’on ne sache pas exactement quelle partie du patrimoine génétique est responsable ne doit pas nous faire classer ces cas inconnus dans les cas de dérive.
Cette distinction est très importante pour comprendre l’impact de ces deux forces évolutives sur les populations et sur leur survie à long terme. En effet, la sélection va avoir tendance à éliminer certains gènes et à en privilégier d’autres modifiant ainsi le patrimoine génétique d’une population de façon NON aléatoire alors que la dérive elle est aveugle et élimine les gènes de façon aléatoire. La sélection va permettre à une population d’être (en gros) mieux adaptée à son environnement alors que la dérive peut potentiellement éliminer aussi des individus adaptés.
Le manque de connaissance sur certains mécanismes de sélection et de déterminisme génétique ajoute de l’aléatoire sur les conclusions mais cet aléatoire pourra (peut-être) être réduit. Ce n’est pas le cas pour l’aléatoire lié à la dérive.
Deux jumeaux blancs peuvent avoir des chances de survie différentes notamment si l’un se ramassent une tuile sur la tête par exemple (dérive), ils peuvent avoir des risques de coups de soleils plus grands que des amis noirs (sélection). Il est peu probable qu’on trouve un jour le gène qui fait éviter les tuiles.
J’espère que j’ai aidé à la compréhension plutôt que l’inverse…
Merci pour ce message et toutes ces précisions à propos des implications différentes de la sélection et de la dérive génétique. Juste quelques petites remarques en réponse à ce message :
– tout d’abord, je ne faisais pas particulièrement référence à ces phénomènes génétiques dans ma réponse sur le hasard, mais plutôt au traitement du hasard en science en général, en m’appuyant sur des exemples démographiques. Dans ce contexte, votre réponse est intéressante car elle illustre bien ce rôle clé de l’observation dans l’interprétation de l’aléa : contrairement au démographe, qui mesure des taux vitaux, le généticien «observe» des génotypes, «mesure» des valeurs sélectives. Il a donc défini son hasard à lui (la dérive génétique) en fonction de ce qui échappe à ses observations.
– Lorsque l’on dit que «la dérive est aveugle et élimine les gènes de façon aléatoire», il s’agit d’un raccourci de langage. En réalité, la dérive n’élimine pas les allèles, c’est l’élimination des allèles (et + généralement leur fluctuation) qui constitue la dérive. En outre, il est incorrect de dire que «la dérive élimine les individus». Ici, vous confondez cause et conséquence. La disparition (ou la non reproduction) de certains individus indépendamment de leur génotype est un facteur qui conduit à la fluctuation des fréquences alléliques que l’on nomme dérive génétique (et non l’inverse).
– Je ne pense pas que du point de vue du généticien (et donc du génotype), l’on puisse dire que 2 jumeaux ont des chances de survie différentes si l’un se prend une tuile sur la tête. C’est la réalisation qui est différente, non la chance. Mais votre exemple illustre vraiment bien les relations fortes qui lient stochasticité démographique (l’un des thèmes développés dans ce Regard) et dérive génétique : les fluctuations des tailles de petites populations et de leurs fréquences alléliques sont généralement les conséquences des mêmes phénomènes (environnementaux) mal décrits et donc considérés comme imprédictibles.