
Influence du calendrier reproductif des femelles babouins chacma sur les conflits mères-enfants
Jules Dezeure
Email: jules.dezeure@etu.umontpellier.fr
1093-1317 Route de Mende, 34090, Montpellier
(Version PDF)
Résumé
Certains primates, dont les Hommes, sont capables de se reproduire tout au long de l’année, bien que vivant dans une grande gamme de latitudes. Cette reproduction non-saisonnière pourrait avoir évolué en réponse à l’absence de variations de succès reproducteur au cours de l’année, découlant probablement d’une habilité à tamponner les fluctuations saisonnières environnementales, via le stockage d’énergie ou un régime omnivore par exemple.
Ici, nous avons utilisé une combinaison de données de long-terme et d’observations comportementales sur une population sauvage de babouins chacmas (Papio ursinus), vivant dans une savane saisonnière en Namibie, pour tester l’adaptabilité de la variation saisonnière des fréquences des naissances, en quantifiant des coûts de fitness associés à des déviations avec les timings de naissance optimaux dans le cycle annuel.
Premièrement, nous avons caractérisé leur reproduction non-saisonnière, dans la mesure où les femelles conçoivent et donnent naissance tout au long de l’année, malgré la saisonnalité importante de leur environnement. Il existe toutefois un léger pic de naissances dans cette population, avec 43 % d’entre elles qui ont lieu en 4 mois, entre Septembre et Décembre. De plus, nous avons également révélé des coûts en termes de fitness de cette reproduction non-saisonnière, à la fois pour les mères et pour leurs petits. En effet, certains timings de naissance minimisent les intervalles entre naissance des femelles (en Septembre), alors que d’autres minimisent la mortalité des jeunes jusqu’à la fin du sevrage (en Décembre). Le pic de naissances observé est donc adaptatif et le résultat du compromis des femelles, visant à maximiser leur rythme reproductif tout en minimisant la mortalité de leurs petits. Grâce à des observations comportementales de terrain sur des juvéniles de moins de 2 ans, nous avons pu mettre en évidence le fait que ce conflit mères-jeunes sur le timing des naissances se traduit au niveau comportemental. La fréquence des crises de colère – la manifestation typique des jeunes primates en conflit avec leurs mères – augmente pour les jeunes nés dans le timing optimal des mères. Ceci peut s’expliquer par un sevrage précoce des jeunes nés à ce moment de l’année, en lien avec un début de sevrage favorisé car synchronisé avec le pic saisonnier de nourriture. La fin de leur sevrage se déroule quant à lui au milieu de la saison sèche, provoquant une augmentation des crises de colère à cette période de faible disponibilité alimentaire. Enfin, le rang social de mères est le seul prédicteur individuel affectant la résolution de ce conflit : les femelles dominantes donnent plus souvent naissance dans le timing optimal pour les mères.
Ainsi, cette étude met en exergue des coûts importants sur le succès reproducteur de se reproduire de façon non-saisonnière, et, met de plus en lumière, pour la première fois, un conflit mère-jeune évolutif sur le timing optimal des naissances, ainsi que sur la variation individuelle des stratégies de résolution de ce conflit. Ces résultats ouvrent de nouvelles perspectives dans la compréhension de l’évolution de la phénologie reproductive des vertébrés.
Justification
La saisonnalité environnementale se caractérise par une fluctuation annuelle de paramètres climatiques, telles que la température ou les précipitations. Celle-ci conduit à une saisonnalité des ressources disponibles, pour un environnement et pour une espèce donnée (Boyce, 1979). Chez les primates, essentiellement frugivores et herbivores, la phénologie des arbres tient un rôle primordial dans cette saisonnalité des ressources disponibles (Di Bitetti & Janson, 2000; van Schaik & van Noordwijk, 1985). La reproduction est une composante essentielle de la valeur sélective d’un individu, et représente, de la conception au sevrage, la phase la plus énergivore dans la vie d’une femelle mammifère adulte (Emery Thompson, 2013; Garcia, 2014). On s’attend donc à observer, comme adaptation de la reproduction à cette saisonnalité climatique et écologique, l’émergence d’une saisonnalité reproductive, se caractérisant par un regroupement temporel d’évènements reproductifs (Lindburg, 1987).
Toutefois, de nombreuses espèces, et notamment l’Homme (Homo sapiens), sont capables de se reproduire tout au long de l’année. Cette reproduction non-saisonnière pourrait avoir joué un rôle-clé dans leur succès écologique. Il est souvent pensé que leur habilité à tamponner les fluctuations environnementales saisonnières est à l’origine de l’évolution de cette non-saisonnalité reproductive, via leur capacité à extraire des ressources en toute saison, et à stocker des quantifiées importantes d’énergie (Bercovitch & Harding, 1993; Brockman & van Schaik, 2005). On s’attend ainsi à ce que les espèces non-saisonnières n’aient aucun coût en terme de valeurs sélectives à se reproduire tout au long de l’année. Toutefois, bien que les coûts de certains timings reproductifs soient bien connus et quantifiés chez de nombreuses espèces saisonnières (SmaIl & Glenn Smith, 1986; Tecot, 2010), ce n’est pas le cas chez des espèces non-saisonnières. Cette étude vise donc à combler cette lacune de connaissances, avec une approche originale, visant à distinguer la saisonnalité de la reproduction optimale en prenant la perspective à la fois des mères et des jeunes. De plus, les déterminants des relations conflictuels entre mère-jeune pendant le sevrage restent méconnus chez les primates sauvages (Maestripieri, 2002).
Méthodes
Le babouin chacma est un primate social vivant dans des groupes multimâles multifemelles, et les femelles sont philopatriques (Barrett & Henzi, 2008; Fischer et al., 2019). Il présente une saisonnalité reproductive étonnante : bien que vivant dans des zones de savanes plus ou moins désertiques fortement saisonnières, il est capable de se reproduire tout au long de l’année (Swedell, 2011). Cette saisonnalité reproductive paradoxale et incomprise en fait un sujet d’étude de choix.
Le Tsaobis Baboon Project (https://www.zsl.org/science/research/baboon), basé dans le Tsaobis Nature Park en Namibie, suit 2 à 3 troupes habituées depuis 2000. À ce jour, il existe trois troupes de babouins habituées à Tsaobis : deux troupes d’environ 70 individus, et une de seulement 20 individus. Chaque individu (même les juvéniles) est identifié et connu, et des suivis journaliers sont effectués de 3 à 6 mois chaque année en Namibie. Le paysage de Tsaobis est constitué de montagnes rocheuses escarpées débouchant vers des plaines alluviales, et coupées au milieu du parc par une rivière éphémère, le lit de rivière de la Swakop (Figure 1).

Figure 1 : Jeunes babouins (de moins de deux ans) observés à Tsaobis Nature Park en Namibie.
Des données de long-terme d’histoire de vie collectées entre 2005 et 2020 nous ont permis de rassembler 215 naissances, 241 conceptions et 171 reprises de cycle (événement reproductif arrive juste après la lactation, quand la femelle démarre un cycle reproducteur, ou non), et d’établir ainsi la saisonnalité de la reproduction de cette population. Des données satellitaires ont été utilisées pour caractériser la saisonnalité environnementale de Tsaobis, plus précisément les variations de pluie, et de ressources disponibles pour les babouins (en utilisant le NDVI, ‘Normalized Difference Vegtation Index’, comme proxy).
Après avoir caractérisé la saisonnalité environnementale et reproductive à Tsaobis, nous avons quantifié les coûts à se reproduire tout au long de l’année en considérant deux indicateurs de succès reproductifs, dans la perspective à la fois des mères et des jeunes, à savoir respectivement les intervalles entre naissances (en jour), et la mortalité des jeunes jusqu’à la fin du sevrage.
Enfin, des observations de terrain effectuées sur 69 jeunes de moins de 2 ans (moyenne ± écart-type = 17.05 ± 7.78 heures d’observation par juvéniles) durant 4 saisons de terrain, en 2006, 2017, 2018 et 2019, ont été utilisées pour quantifier l’investissement maternel et les conflits mère-jeune jusqu’au sevrage (voir Figure 1). Des observations focales de 20 minutes ont été réalisées, durant lesquelles l’activité de l’individu focal (ici le jeune de moins de 2 ans) était noté, en addition de comportements liés à des crises de colère avec sa mère (geck, cris stridents, se jeter au sol, en tournoyant la queue, etc). Plus précisément, nous nous demandons si les mères sont capables de limiter l’effet des fluctuations saisonnières de l’environnement sur le développement des jeunes par leur ajustement d’investissement maternel (fréquence de tétée et fréquence de transport sur la mère). De plus, nous nous attendons à ce que les jeunes nés hors du timing optimal de naissance soient plus colériques, dans la mesure où ils ne sont pas sevrés au moment optimal de l’année en terme de nourriture disponible.
Résultats et discussion
Comme attendu au vu d’autres populations de babouins (Swedell, 2011), les babouins chacma de Tsaobis sont non-saisonniers, c’est à dire qu’ils se reproduisent, donnent naissance et peuvent commencer un nouveau cycle reproducteur tout au long de l’année (Figure 2). L’environnement est hautement saisonnier, avec une saison des pluies entre Janvier et Avril qui est suivi par un pic de nourriture disponible pour les babouins de Mars à Mai (Figure 2). Un léger pic de naissance est observé, avec 43% des naissances ayant eu lieu sur une période de 4 mois, de Septembre à Décembre, c’est à dire à la fin de la saison sèche, juste avant la saison des pluies. Ce genre de patrons se retrouve chez de nombreuses population de babouins, non-saisonniers mais avec des variations saisonnières de fréquence de naissance, et un pic de naissance précédent la saison des pluies (Cheney et al., 2004; Lycett, Weingrill, & Henzi, 1999).
-
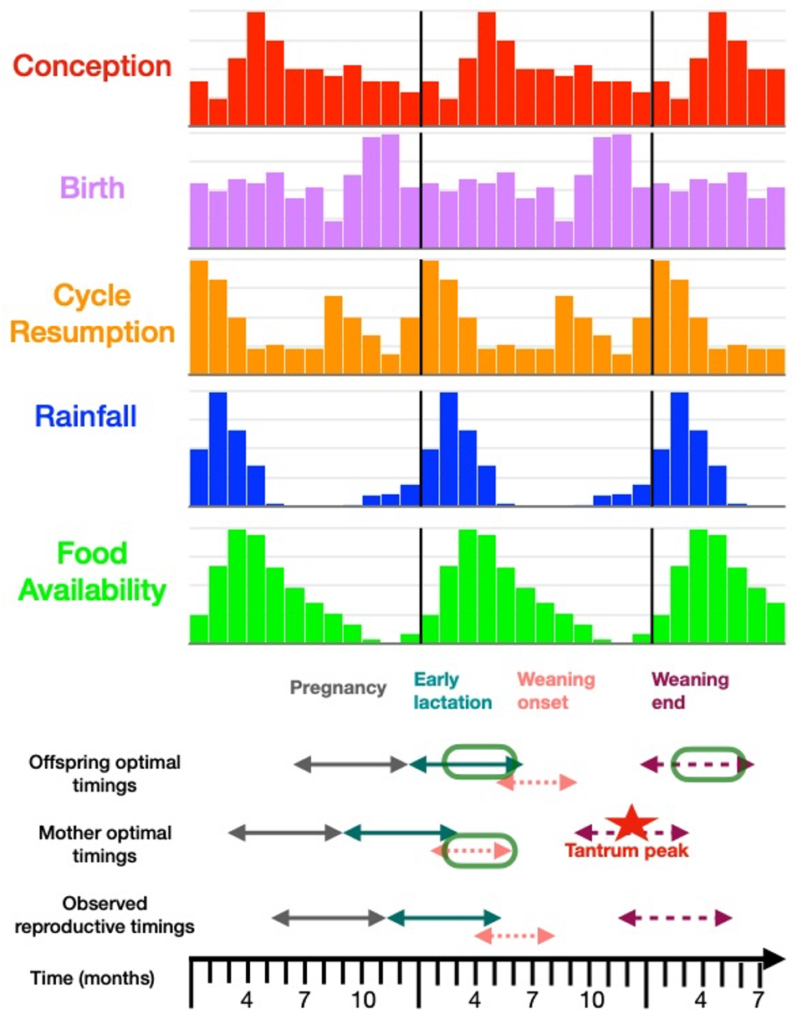
- Figure 2 : Résumé du calendrier reproductif des femelles babouins de Tsaobis, en relation avec les fluctuations saisonnières de pluie et de ressources disponibles.
-
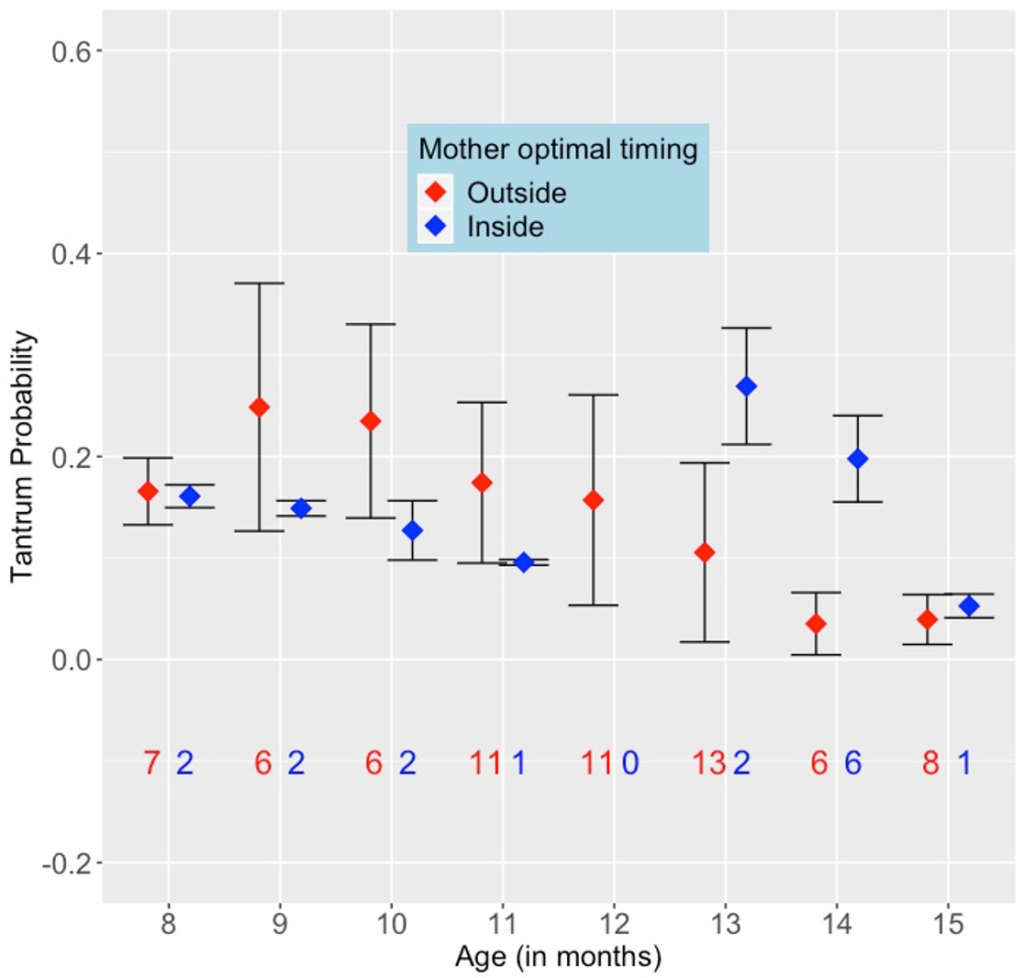
- Figure 3 : Importance du timing de naissance sur les fréquences de crise de colère des jeunes, selon leur âge. Les fréquences de colère en fonction de l’âge, en mois, sont représentées ici pour les fourchettes âges où des données étaient disponibles sur des jeunes nés à la fois hors et durant le timing optimal des mères. Les points rouges représentent les valeurs moyennes par âge du jeune pour ceux nés hors du timing optimal maternel, alors que les points bleus représentent celles pour les jeunes dans ce timing optimal, c’est à dire entre le 23 juillet et le 23 septembre. Les barres verticales indiquent les écart-types associés à chaque valeur moyenne. Le nombre de jeunes observées pour chaque âge né en dehors ou dans le timing optimal maternel est indiqué respectivement en rouge et en bleu.
Étonnamment, nous avons trouvé des coûts importants à cette reproduction non-saisonnière sur nos deux indicateurs de valeurs sélectives : les intervalles entre naissances des mères sont plus courts pour celles ayant donné naissance en Août-Septembre ; alors que la mortalité des jeunes avant sevrage est minimisée pour ceux nés en Décembre. Ainsi, nous avons révélé, pour l’une des première fois, des coûts en terme de valeurs sélectives à donner naissance à certains moments de l’année pour une espèce à reproduction pourtant non-saisonnière. Nous avons donc de plus révélé deux timing de naissance optimaux, séparés de 3 mois, un timing maternel (en Septembre), et un timing juvénile (en Décembre). Le pic de naissance observé qui se retrouve entre ces deux optimums est donc adaptatif, et est le résultat d’un compromis des femelles cherchant à minimiser leur intervalle entre naissance tout en maximisant la survie de leurs jeunes (Figure 2).
Enfin, et contrairement à nos hypothèses, nous n’avons pas trouvé d’ajustement des investissement maternels selon les timings de naissance. C’est à dire que les fréquences des tétées et de transport sur la mère diminuent avec l’âge du jeune, mais ne varient pas selon sa date de naissance. Toutefois, nous avons trouvé un effet significatif de la date de naissance, en contrôlant pour l’âge du jeune, sur la fréquence des colères : les jeunes nés dans l’optimal de timing maternel (en Septembre) ont tendance à être plus colérique que les autres, surtout lorsqu’ils ont plus d’un an (Figure 3).
Le timing optimal pour les jeunes permet de synchroniser deux périodes critiques pour les femelles et jeunes avec le pic saisonnier de nourriture : le début de la lactation et la fin du sevrage (Figure 2). Le timing optimal pour les mères permet quant à lui de synchroniser le début de sevrage, quand le jeune a environ 5-6 mois, avec ce pic de nourriture (Figure 2). C’est un âge charnière dans le développement des jeunes babouins (Altmann, 1980; Rhine, Wasser, & Norton, 1988), et de ce fait, les mères peuvent sevrer leur jeune précocement, et se reproduire par la suite plus rapidement. Néanmoins, ceci vient avec un coût pour les jeunes, qui se retrouvent avec une fin de sevrage en pleine saison sèche, période de faible abondance de nourriture. Ceci augmente donc leur chance de mortalité, et augmente la fréquence de leur crises de colère, qui sont ici utilisées comme signal honnête de requête d’investissement maternel supplémentaire (Barrett & Henzi, 2000; Hauser & Fairbanks, 1988). De plus, nous avons également pu montrer que les femelles dominantes donnent plus souvent naissance que les subordonnées dans ce timing, ce qui suggèrent qu’il est également coûteux pour certaines femelles. En effet, le début de la lactation est une période énergétiquement délicate pour les femelles (Lee, 1996), et ainsi, seules les femelles dominantes parviennent à limiter ces coûts énergétiques dans cette population.
Les babouins chacma de Tsaobis se reproduisent de façon non-saisonnière, alors que cela est coûteux : d’autres pressions de sélection, en lien avec des facteurs sociaux ou l’imprévisibilité de l’environnement par exemple, doivent donc être à l’origine de l’absence de reproduction saisonnière.
Références
Altmann, J. (1980). Baboon Mothers and Infants. The University of Chicago Press.
Barrett, L., & Henzi, S. P. (2000). Are baboon infants Sir Phillip Sydney’s offspring? Ethology. https://doi.org/10.1046/j.1439-0310.2000.00577.x
Barrett, L., & Henzi, S. P. (2008, May 20). Baboons. Current Biology, Vol. 18. https://doi.org/10.1016/j.cub.2008.02.074
Bercovitch, F. B., & Harding, R. S. O. (1993). Annual Birth Patterns of Savanna Baboons (Papio cynocephalus anubis) over a Ten-Year Period at Gilgil, Kenya. In Folia Primatol (Vol. 61).
Boyce, M. S. (1979). Seasonality and Patterns of Natural Selection for Life Histories. The American Naturalist. https://doi.org/10.1086/283503
Brockman, D., & van Schaik, C. (2005). Seasonality and reproductive function. In Cambridge University Press (Ed.), Seasonality in Primates: Studies of Living and Extinct Human and Non-Human Primates. Retrieved from https://www.researchgate.net/publication/230778872
Cheney, D. L., Seyfarth, R. M., Fischer, J., Beehner, J., Bergman, T., Johnson, S. E., … Silk, J. B. (2004). Factors Affecting Reproduction and Mortality Among Baboons in the Okavango Delta, Botswana. International Journal of Primatology, 25(2), 1138–479986.
Di Bitetti, M. S., & Janson, C. H. (2000). When will the stork arrive? Patterns of birth seasonality in neotropical primates. American Journal of Primatology. https://doi.org/10.1002/(SICI)1098-2345(200002)50:2<109::AID-AJP2>3.0.CO;2-W
Emery Thompson, M. (2013). Comparative Reproductive Energetics of Human and Nonhuman Primates. Annual Review of Anthropology, 42(1), 287–304. https://doi.org/10.1146/annurev-anthro-092412-155530
Fischer, J., Higham, J. P., Alberts, S. C., Barrett, L., Beehner, J. C., Bergman, T. J., … Zinner, D. (2019). The Natural History of Model Organisms: Insights into the evolution of social systems and species from baboon studies. ELife, 1960, 1–16.
Garcia, C. (2014). Cost of reproduction in female non-human primates: an anthropological perspective. BMSAP. https://doi.org/10.1007/s13219-014-0106-1
Hauser, M. D., & Fairbanks, L. A. (1988). Mother-offspring conflict in vervet monkeys: variation in response to ecological conditions. Animal Behaviour. https://doi.org/10.1016/S0003-3472(88)80163-5
Lee, P. C. (1996). The Meanings of Weaning: Growth, Lactation, and Life History. Evolutionary Anthropology.
Lindburg, D. (1987). Seasonality of Reproduction in Primates. In G. Mitchell & J. Erwin (Eds.), Comparative Primate Biology (Alan R. Li, pp. 167–218). New York.
Lycett, J. E. ., Weingrill, T. ., & Henzi, S. . (1999). Birth patterns in the Drakensberg Mountain baboons (Papio cynocephalus ursinus). South African Journal of Science, 95(8), 354–356.
Maestripieri, D. (2002). Parent – Offspring Conflict in Primates. International Journal of Primatology, 23(4), 2005–2007. https://doi.org/10.1023/A:1015537201184
Rhine, R. J., Wasser, S. K., & Norton, G. W. (1988). Eight-Year Study of Social and Ecological Correlates of Mortality Among Immature Baboons of Mikumi National Park, Tanzania. American Journal of Primatology, 16, 199–212.
SmaIl, M. F., & Glenn Smith, D. (1986). The Influence of Birth Timing upon Infant Growth and Survival in Captive Rhesus Macaques (Macaca mulatta). International Journal of Primatology, 7(3).
Swedell, L. (2011). African papionins: diversity of social organization and ecological flexibility. In Primates in perspective (pp. 241–277). Retrieved from https://www.researchgate.net/publication/285789215
Tecot, S. R. (2010). It’s All in the Timing: Birth Seasonality and Infant Survival in Eulemur rubriventer. International Journal of Primatology. https://doi.org/10.1007/s10764-010-9423-5
van Schaik, C. P., & van Noordwijk, M. A. (1985). Interannual variability in fruit abundance and the reproductive seasonality in Sumatran Long-tailed macaques (Macaca fascicularis). J. Zool., Lond. ( A ), 206, 533–549.
Commentaires récents