La Société Française d’Ecologie et d’Evolution (SFE2) vous propose ce regard de Régis Céréghino, Professeur à l’Université Paul Sabatier Toulouse 3, sur la diversité des stratégies écologiques des espèces, confrontées aux changements globaux.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions sur les forums de discussion qui suivent les articles; les auteurs vous répondront.
———-
Quelle diversité des stratégies écologiques,
face aux changements globaux ?
par Régis Céréghino,
Professeur à l’Université Paul Sabatier Toulouse 3. EcoLab, Laboratoire Ecologie Fonctionnelle et Environnement UMR5245 CNRS-UPS-INPT. regis.cereghino@univ-tlse3.fr
Article édité par Anne Teyssèdre
——-
Mots clés : Traits fonctionnels, communautés, diversité biologique, espace des traits,
changements globaux, adaptation, résilience, extinctions.
——–
- Introduction
- L’espace des traits
- Contraintes sur les stratégies écologiques
- Conclusion
- Bibliographie
- Regards connexes
- Forum de discussion sur ce regard
——-
Introduction
L’un des principaux objectifs de l’écologie est de comprendre l’incidence des changements environnementaux sur la diversité biologique et le fonctionnement des écosystèmes. Pour atteindre cet objectif, les écologues ont besoin de modèles mécanistes qui relient les organismes entre eux et avec leur environnement. Les traits fonctionnels, attributs biologiques, physiologiques et écologiques des organismes, déterminent la façon dont ces derniers interagissent avec leur environnement biologique et physique (i.e. biotique et abiotique) (cf. Fig. 1). Il s’agit par exemple de la taille et de la forme du corps, du cycle de vie, du mode de respiration, du régime alimentaire, des modes de locomotion et de dispersion, des habitats préférentiels, des moyens de résister aux prédateurs ou à des perturbations récurrentes. Parmi les traits fonctionnels, on peut distinguer les traits d’histoire de vie (ex. modalité de croissance, âge à maturité, taille de la progéniture, durée de vie) liés à la reproduction et à la descendance des individus, et par extension à la démographie des populations.

Figure 1. Photos de quelques arthropodes représentant une diversité de traits fonctionnels et de rôles écologiques au sein des écosystèmes : a) Araignée salticide et sa proie (© Olivier Dézerald), b) Fourmis coupeuses de feuilles (© Bruno Corbara), c) Bourdon des champs sur une anémone (© Claire Céréghino Macret), d) Larve de charançon se nourrissant de feuilles de broméliacée (© Bruno Corbara).
Tandis que les conditions et les ressources environnementales définissent les potentiels de colonisation et de viabilité pour les populations, les traits fonctionnels prédisent les performances des organismes dans ces espaces multidimensionnels. Ils sont donc plus informatifs que la seule identité des espèces pour déchiffrer les mécanismes qui sous-tendent la dynamique de la biodiversité. En s’affranchissant des différences taxonomiques entre régions biogéographiques et types d’écosystèmes, ils permettent en outre d’établir des généralités sur les relations environnement-biodiversité-fonctionnement écologique, plutôt que de collectionner des études de cas.
Il existerait 8 millions d’espèces actuelles sur Terre, dont 1.8 millions seulement sont connues (Mora et al., 2011), et près d’un million d’espèces d’animaux et de plantes seraient menacées d’extinction par les activités humaines (www.IPBES.net). La richesse en espèces ne s’accompagne cependant pas d’une richesse équivalente de traits fonctionnels (Céreghino et al., 2018). A priori, les faunes et flores d’écosystèmes ou régions entiers devraient être d’autant plus vulnérables que la diversité fonctionnelle des espèces est faible, et qu’un grand nombre d’espèces partagent les mêmes traits de sensibilité aux changements hydrologiques, thermiques, chimiques, etc. (voir les regards n°16 et 80a). Par conséquent, il est urgent d’identifier les combinaisons de traits fonctionnels qui définissent les stratégies écologiques des espèces face aux fluctuations environnementales, et de comprendre les contraintes écologiques et évolutives qui façonnent ces stratégies. Les réponses à ces questions aideront les écologues à prévoir l’impact des changements environnementaux sur les communautés biologiques et les processus écosystémiques qui y sont associés.
L’espace des traits
Les combinaisons de traits fonctionnels qui définissent les stratégies écologiques des plantes et des animaux sont souvent réduites à cinq aspects fondamentaux (Winemiller et al., 2015 ; Pianka et al., 2017) : i) type trophique (sources et moyens pour se procurer de l’énergie), habitat (types, spécialisation), ii) histoire de vie (taille à maturité, taux de croissance, iii) durée du cycle de vie, complexité, fécondité, soins parentaux), iv) type de défense (active, passive, morphologique, chimique), et v) métabolisme. Chez les animaux par exemple, les combinaisons de traits liés aux histoires de vie (ex. taille, croissance, nombre de descendants) et à l’habitat (ex. spécialistes vs généralistes) reflètent la capacité de résilience des populations face à des évènements extrêmes, tandis que les traits trophiques expriment les interactions – ou la dépendance – avec les autres membres de la communauté, les réponses potentielles aux changements de ressources, ou encore le rôle des espèces sur des processus comme la décomposition de la matière organique ou la pollinisation.
Peu de travaux ont permis de comparer la diversité des combinaisons de traits fonctionnels réalisées aux combinaisons théoriquement possibles à l’échelle de faunes et de flores entières, mais tous pointent vers la même conclusion : au regard de leur forte diversité taxonomique, les plantes et les animaux n’ont développé qu’un tout petit nombre de stratégies écologiques fructueuses. Selon Cooke et al. (2019), les 15000 espèces actuelles de mammifères et d’oiseaux ne réalisent que 9% des combinaisons de traits possibles. De la même façon, les 46000 espèces de plantes vasculaires du monde occupent moins de 23% de leur espace de traits potentiels (Diaz et al., 2016), une restriction similaire à celle observée pour les 850 espèces d’invertébrés aquatiques vivant dans les broméliacées à réservoir d’eau des Néotropiques (Céréghino et al., 2018). Des agrégations d’espèces (26% de l’espace fonctionnel potentiel) ont également été observées chez des invertébrés et vertébrés néotropicaux en utilisant la composition élémentaire des tissus (carbone, azote, phosphore) en tant que « traits biochimiques » (González et al. 2017).
Contraintes sur les stratégies écologiques
Dans l’univers des stratégies écologiques possibles, c’est-à-dire parmi toutes les combinaisons de traits théoriquement possibles, l’espace des traits effectivement réalisé par un pool d’espèces local à global est limité par des compromis physiologiques entre les traits, ainsi que par des contraintes évolutives et écologiques.
Premièrement, les organismes ne peuvent optimiser simultanément toutes leurs performances biologiques et physiologiques. Des compromis limitent donc les combinaisons de traits possibles. Un exemple bien connu est la relation entre la forme et la taille du corps (Raup, 1966) et ses conséquences sur la physiologie des invertébrés. Les invertébrés aquatiques de forme cylindrique ont un rapport surface externe / volume faible, qui autorise une respiration au travers du tégument (sans intervention de surface supplémentaire spécialisée, comme celle des branchies ou des poumons), mais au-delà d’une certaine taille corporelle les organes profonds ne sont plus approvisionnés et la respiration par simple diffusion n’est plus possible (Barnes et al., 2009).
Deuxièmement, des contraintes évolutives conduisant à un conservatisme phylogénétique peuvent restreindre l’espace des traits. Au fil du temps, le conservatisme phylogénétique préserve de nombreux traits chez les espèces appartenant aux mêmes familles ou genres (Pianka et al., 2017). Les espèces qui sont proches sur l’arbre du vivant se concentrent alors dans l’espace des traits fonctionnels ; elles ont des stratégies écologiques proches (cf. Fig. 2 ci-dessous). Un bon exemple est celui des invertébrés d’eau douce d’Europe (Usseglio-Polatera et al., 2000), d’Amérique du Nord (Poff et al., 2006) et des Néotropiques (Céréghino et al., 2018), où les traits fonctionnels agrègent les espèces par famille puis les familles par ordres.
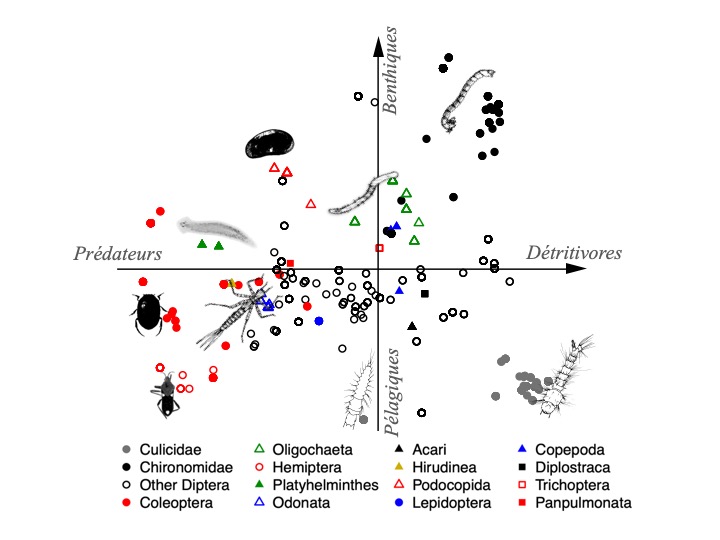
Figure 2. Exemple de distribution d’espèces dans un espace de traits fonctionnels à deux dimensions (trophique : prédateurs à détritivores ; habitat préférentiel : pélagique à benthique), et de conservatisme phylogénétique des traits fonctionnels chez les invertébrés néotropicaux vivant dans les broméliacées à réservoir (d’après Céréghino et al., 2018). Chaque point correspond à une espèce ; les différents marqueurs indiquent les familles, ordres or phylums auxquels elles appartiennent. Les espèces se distribuent dans cet espace selon leurs traits fonctionnels, et tendent à s’agréger par groupe taxonomique. Quelques illustrations représentent les principaux groupes (ex. odonates, culicides…).
On peut conclure que le conservatisme phylogénétique est un mécanisme répandu de diversification des stratégies écologiques chez ces invertébrés. Les écarts majeurs de stratégies écologiques se situent ici entre les insectes (aquatiques) et les autres ‘invertébrés’, puis entre les différentes lignées d’invertébrés hors insectes (crustacés, annélides, etc.). Cela n’est pas surprenant dans la mesure où les grandes lignées évolutives naissent d’innovations biologiques.
Par ailleurs, la convergence évolutive tend aussi à concentrer des espèces phylogénétiquement distantes dans l’espace des traits. Dans ce cas, des espèces non apparentées adoptent des traits et stratégies similaires en réponse à une sélection par les contraintes environnementales, réduisant ainsi l’occupation globale de l’espace fonctionnel. La forme hydrodynamique et les « nageoires » des poissons, des cétacés et des manchots en est un exemple.
Troisièmement, l’habitat agit comme un filtre pour les stratégies écologiques, en éliminant les espèces aux stratégies non viables au regard des contraintes environnementales. Pour les communautés de ces écosystèmes, les stratégies non viables se traduisent par des régions vides dans l’espace des traits fonctionnels réalisés. L’échelle spatiale (habitat, site, région, continent…) est ici implicite, car différent filtres (barrières de dispersion, ressources, compétition, prédation) sont susceptibles d’agir à différentes échelles.
Conclusion
Si plusieurs facteurs peuvent contraindre les stratégies écologiques des espèces, il semble que celles-ci soient surtout liées au conservatisme des traits et donc à la position des espèces sur l’arbre phylogénétique (Pianka et al., 2017 ; Céréghino et al., 2018). Ceci suggère que l’apparition de nouvelles combinaisons de traits est très lente à l’échelle des temps évolutifs – et beaucoup trop lente pour que le vivant puisse faire face à des changements rapides dont les causes ne se dissipent pas. Sur la base des probabilités d’extinction des espèces, une étude récente suggère que le nombre très limité de stratégies viables chez les mammifères et les oiseaux devrait diminuer encore au cours du siècle à venir, au profit d’espèces généralistes, de petite taille, et à forte fécondité (Cooke et al., 2019, voir aussi les regards n°16, 45 et 80a).
Cette tendance rappelle l’extinction de la mégafaune du Pléistocène entre -50 000 et -15000 ans dans le passé (très probablement causée par la conjonction de la colonisation de continents et îles par notre espèce et de changements climatiques), et ses conséquences significatives sur les interactions biologiques et flux de nutriments (e.g. Doughty et al. 2016, et voir le regard n°80b). Ou encore, on estime que le transport de phosphore depuis les océans vers les terres par les oiseaux de mer et les poissons anadromes a été réduit de 96% au cours des derniers siècles, du fait de l’intensification des activités humaines – chasse à la baleine, décimation de colonies de phoques et oiseaux de mer, intensification de la pêche, barrages, etc. (Doughty et al., op. cit.).
Les traits fonctionnels et les stratégies écologiques qui en découlent apportent donc un éclairage différent sur la biodiversité : les espèces n’ont pas qu’une valeur patrimoniale, et toutes sont impliquées directement ou indirectement dans des processus écosystémiques dont l’Homme dépend de façon ultime. Si le message est bien connu des scientifiques, il mériterait sans doute un plus fort relais auprès des décideurs, des gestionnaires de l’environnement et du public car il définit mieux notre dépendance aux multiples facettes de la diversité biologique.
Bibliographie
Barnes R.S.K., P.P. Calow P.J.W., Olive D.W., Golding & J.I. Spicer, 2009. The invertebrates: a synthesis, 3rd Edition. Wiley-Blackwell.
Céréghino R., V. Pillar, D. Srivastava et al., 2018. Constraints on the functional trait space of aquatic invertebrates in bromeliads. Functional Ecology 32, 2435-2447.
Cooke R.S.C, F. Eigenbrod & A.E. Bates, 2019. Projected losses of global mammal and bird ecological strategies. Nature Communications 10, 2279.
Díaz S., J. Kattge, J.H.C. Cornelissen, I.J. Wright, S. Lavorel, S. Dray, (…) & L.D. Gorné, 2016. The global spectrum of plant form and function. Nature 529, 167–171.
Doughty C.E., J. Roman, S. Faurby, A.Wolf, A. Haque, E.S. Bakker, Y. Malhi, J.B. Dunning Jr & J.C. Svenning, 2016. Global nutrient transport in a world of giants. Proceedings of the Natural Academy of Sciences USA 113, 868–873.
González A.L., O. Dézerald, P.A. Marquet, G.Q. Romero & D. Srivastava, 2017. The multidimensional stoichiometric niche. Frontiers in Ecology and Evolution 5, 110.
Mora C., D.P. Tittensor, S. Adl, A.G.B. Simpson & B. Worm, 2011. How Many Species Are There on Earth and in the Ocean? PLoS Biology 9, e1001127
Pianka E.R., L.J. Vitt, N. Pelegrin, D.B. Fitzgerald & K.O. Winemiller, 2017. Toward a periodic table of niches, or exploring the lizard niche hypervolume. The American Naturalist, 190 601-616.
Poff N.L., J.D. Olden, N.K.M. Viera, D.S. Finn, M.P. Simmons & B.C. Kondratieff, 2006. Functional trait niches of North American lotic insects: traits based ecological applications in light of phylogenetic relationships. Journal of the North American Benthological Society 25, 730–755.
Raup D.M., 1966. Geometrical analysis of shell coiling: general problems. Journal of Palaeontology 40, 1178–1190.
Usseglio-Polatera P., M. Bournaud, P. Richoux & H. Tachet, 2000. Biological and ecological traits of benthic freshwater macroinvertebrates: relationships and definition of groups with similar traits. Freshwater Biology 43, 175–205.
Winemiller K.O., D.B. Fitzgerald, L. Bower & E.R. Pianka, 2015. Functional traits, convergent evolution, and periodic tables of niches. Ecology Letters 18, 737–751.
—-
Regards connexes :
Clavel J., 2011. L’homogénéisation biotique, une réponse aux changements globaux. Regards et débats sur la biodiversité, SFE, Regard n°16, avril 2011.
Julliard R. et F. Jiguet, 2011. Les oiseaux et la biodiversité face au changement climatique. Regards et débats sur la biodiversité, SFE, Regard n°22, oct. 2011.
Lebreton J.D., 2013. Biodiversité et dynamique des populations. Regards et débats sur la biodiversité, SFE, Regard n°45, avril 2013.
Teyssèdre A., 2018a. Quelles réponses des espèces et communautés aux changements globaux? Regards et débats sur la biodiversité, SFE, Regard n°80a, juin 2018.
Teyssèdre A., 2018b. La dynamique des mammifères face aux changements globaux. Regards et débats sur la biodiversité, SFE, Regard n°80b, juin 2018.
Article édité et mis en ligne par Anne Teyssèdre.
——
Quel plaisir à la lecture de ce regard ! Clair et concis, tout en s’appuyant sur des exemples précis, sur un sujet brûlant d’actualité. Je vais m’empresser de lire votre article de 2018 dans FUNCTIONAL ECOLOGY…
Bonjour Régis,
et merci pour ce regard sur la diversité des stratégies écologiques des invertébrés d’eau douce.
Pour prolonger ces recherches, avez-vous -toi ou d’autres membres de cette équipe- exploré la variation des traits fonctionnels (au sein des communautés considérées) en réponse aux actuels changements globaux ? C’est-à-dire, par exemple : comment varient ces traits -dans chaque groupe fonctionnel, voire dans chaque espèce ou taxon- avec le degré de transformation ou de fragmentation des habitats, ou avec les variations locales du climat?
[Y a-t-il une augmentation de la variance des traits en réponse à la perturbation des habitats ? Une tendance générale vers certaine(s) direction(s)? Une augmentation des traits ‘généralistes’ vs spécialisés?..]
Bien cordialement, Anne
Réponse à Anne:
Nous avons manipulé la fréquence et l’intensité des précipitations qui entrent dans les broméliacées dans 7 sites distribués entre Puerto Rico et l’Argentine, aussi bien dans le sens de précipitations plus fréquentes et plus intenses que moins fréquentes et moins intenses.
Les traitements de sécheresse ont eu les effets les plus significatifs sur la diversité fonctionnelle. Cependant, les sites qui ont été les plus affectés (perte de traits fonctionnels uniques au sein des communautés) sont ceux qui présentaient naturellement les pools d’espèces les plus pauvres (effet d’isolation dans les Caraïbes, limites de distribution des broméliacées et de leur faune en Argentine).
Cela suggère d’une part que la diversité spécifique joue un rôle d’assurance pour la stabilité fonctionnelle des communautés, et d’autre part que la biogéographie est une composante importante de la réponse fonctionnelle des communautés au changement climatique. Les articles sont en cours de rédaction…. A suivre donc !
Bonjour,
Régis C. m’informe qu’étant en mission en Guyane il ne peut répondre ces jours-ci aux questions et commentaires sur son article/regard en ligne. Il pourra répondre après le 15 octobre. Donc patience!
Bien cordialement à tous, Anne