
La Société Française d’Ecologie (SFE) vous propose le regard de Joanne Clavel sur l’homogénéisation des communautés vivante en réponse aux changements globaux.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
L’homogénéisation biotique :
une réponse aux changements globaux
Joanne CLavel
Département Arts et Sciences de la communication, Faculté de Philosophie et Lettres,
Place du 20 aout, Université de Liège, Belgique
( Fichier PDF )
Regard R16, édité par Anne Teyssèdre
———
Mots clés : homogénéisation biotique, écologie, communautés, dynamique, mécanismes, espèces, niche écologique, interactions, changements globaux, invasions biologiques, homogénéisation fonctionnelle.
———
L’altération des écosystèmes par les activités humaines et la dispersion assistée à grande échelle d’espèces exotiques ont influencé et bouleversé la distribution des espèces. Ces quinze dernières années, de nombreuses études ont révélé un déclin prononcé des espèces animales et végétales spécialistes que ce soit dans des écosystèmes terrestres ou marins. C’est en voulant comprendre pourquoi certaines espèces présentent un plus grand risque de s’éteindre, objet d’étude de la biologie de la conservation, que ces études ont convergé en montrant que le fort degré de spécialisation des espèces augmentait le risque de déclin des populations.
Qu’est ce que la spécialisation d’une espèce ?
Cette notion renvoie à l’un des plus vieux concepts en écologie à savoir celui de « niche écologique ». Ce concept a été développé initialement pour comprendre la coexistence des espèces dans un milieu (Hutchinson 1957). On définit la niche écologique d’une espèce comme sa position au sein d’un écosystème résultant d’une somme de paramètres allant des propriétés physico-chimiques du milieu, aux ressources que l’espèce exploite, à l’ensemble des interactions (positives comme négatives) qu’elle entretient avec les autres espèces. Si on se place à une très grande échelle de temps, les forces évolutives tendent à différencier les niches soit par déplacement des besoins écologiques, soit par variations de la largeur de niche (figure 1). Ce dernier cas conduit à la coexistence d’espèces spécialistes à niche écologique étroite et d’espèces généralistes à niche large, ou plus exactement, des espèces plus ou moins spécialistes ou plus ou moins généralistes (Futuyma and Moreno 1988).
Dans de nombreux cas il semblerait que la spécialisation s’explique par un compromis (trade-off) évolutif entre la capacité d’une espèce à exploiter un gradient de ressource et leur performance sur cette ressource. Cependant, le coût de la généralisation peut aussi théoriquement être lié à un fardeau de mutation délétères et il existe au moins dans le règne des champignons des « super généralistes » qui ne suivent pas l’adage « Qui peut tout faire ne maîtrise rien » (« Jack-of-all-trades is master of none »). Enfin, un lien direct entre le déplacement de caractères et la largeur de niche a été montré dans des relations hôtes/parasites lorsqu’un trait responsable de la spécialisation est également responsable d’un changement d’hôte.
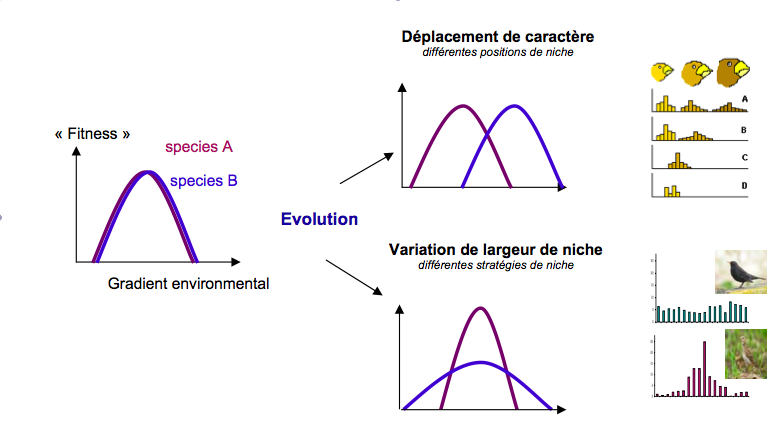
Figure 1. Deux espèces ne peuvent avoir exactement la même niche. L’Evolution mène soit à un déplacement de niche donnant des positions différentes aux espèces, illustré par l’exemple bien connu de variation de taille de bec chez les pinsons de Darwin (Krebs 2001. Ecology, 5th Edn) ou/et elle peut mener à une variation de largeur de niche menant à deux stratégies extrêmes : les espèces spécialistes et généralistes illustrées par la répartition d’abondance de deux passereaux donnant lieu au calcul de l’indice de spécialisation des espèces (SSI) (Julliard et al. 2004).
Alors que les spécialistes sont en déclin, les généralistes semblent à l’inverse bénéficier des changements globaux en colonisant de nouvelles aires de distribution ou en augmentant leur taille de population. Par exemple, il a été montré chez des vertébrés (poissons et oiseaux) que les espèces généralistes ont une forte probabilité d’être introduites et de s’établir (Fisher and Owens 2004). En effet, les généralistes possèdent de plus larges populations tout en étant plus flexibles et peu exigeantes en terme de ressources voire étonnamment innovantes.
Si les changements globaux érodent d’une manière générale la biodiversité, nous venons de montrer que certaines espèces sont moins affectées que d’autres. Cette inégalité de réponse des espèces face aux changements globaux, entraîne presque systématiquement le remplacement d’espèces spécialistes par quelques espèces généralistes. Par exemple, en réponse à la modification des habitats (fragmentation et perturbation liée aux changements temporels des habitats), Devictor et ses collaborateurs ont montré que les communautés d’oiseaux à l’échelle de la France se généralisent (Devictor et al. 2008). Il s’opère donc au sein des écosystèmes dégradés, une « homogénéisation biotique » des communautés de plus en plus composées d’espèces généralistes.
Qu’est ce que l’homogénéisation biotique ?
L’homogénéisation biotique (HB) est historiquement issue de la littérature des espèces invasives. Pendant de longues années les scientifiques se sont principalement intéressés à «l’homogénéisation taxonomique » des communautés, décrite comme une augmentation de similarité dans la composition des communautés par l’invasion des quelques mêmes espèces (figure 2).

Figure 2. Dans les années 2000, le remplacement d’espèces natives par des espèces ubiquistes introduites par les humains était couramment désigné comme la cause principale de l’homogénéisation des communautés à l’échelle mondiale. Tiré de Olden, 2006.
Bien que populaire, la mesure de l’homogénéisation taxonomique mène à des confusions d’interprétations : cette homogénéisation est elle due au gain ou à la perte des mêmes espèces, ou encore à une conjonction des deux phénomènes ? De plus, le concept d’espèce n’a quasi plus de pertinence dans certains groupes taxonomiques. Il existe d’autres niveaux d’intégration de l’Homogénéisation Biotique (Olden et al. 2004) comme l’HB génétique (2) ou l’HB fonctionnelle objet de cet article et que nous allons développer sous l’angle de la spécialisation des espèces.
L’HB fonctionnelle mesure la redistribution d’un ou de plusieurs traits fonctionnels sans considérer l’identité taxonomique des espèces. C’est par ce moyen que l’on peut avoir accès aux mécanismes écologiques qui expliquent la distribution spatiale et la composition des communautés. Pour expliquer l’impact des changements globaux sur les communautés, nous identifions trois mécanismes différents qui aboutissent au remplacement des spécialistes par des généralistes. Premièrement, les perturbations peuvent agir directement comme lors de la destruction d’habitat qui entraine mécaniquement la disparition des espèces spécialisées à cet habitat. Deuxièmement, les espèces généralistes étant plus flexibles et innovantes elles possèdent une plus grande capacité à coloniser de nouvelles niches écologiques. Face au réchauffement climatique, par exemple, les espèces tendent à déplacer leur aire de distribution pour suivre les isoclines de températures. Les espèces généralistes sont capables de réagir plus rapidement car elles tolèrent les modifications d’occupation des sols qui représentent autant de freins aux spécialistes pour remonter vers le nord (Warren et al. 2001).
Enfin, indirectement les changements globaux dégradent la qualité globale des habitats (ressources, qualité des informations disponibles…) et modifient les interactions entre espèces comme la compétition ou la facilitation. Par exemple, l’événement de canicule en France de 2003 a eu tendance à favoriser la reproduction de tous les passereaux, cependant au moment du recrutement l’année suivante, les généralistes ont gagné par compétition et se retrouvaient donc plus abondantes toute proportion gardée que les spécialistes (Julliard et al. 2004).
Quel est impact de l’homogénéisation biotique sur les écosystèmes ?
Comme nous l’avons décrit jusqu’à présent, l’homogénéisation fonctionnelle est corrélée à une dégradation directe ou indirecte de l’écosystème. Cependant, en est-elle le symptôme ou la cause ? C’est là que se trouve tout le cœur du débat futur puisque les recommandations de gestion en terme de conservation ne seront évidemment pas les mêmes selon la réponse. L’homogénéisation fonctionnelle et le nombre d’interrogations qu’elle soulève proposent un nouveau défi à la communauté des écologues. Comment ces changements au sein des communautés altèrent le fonctionnement des écosystèmes ? Qu’en découle-t-il en terme de « services écosystémiques » ?
Sur les trois mécanismes fondamentaux expliquant la relation positive liant diversité biologique et fonctionnement des écosystèmes, deux nous apparaissent comme particulièrement préoccupants dans le contexte d’homogénéisation fonctionnelle. Le premier est celui de facilitation, à savoir l’interaction positive entre deux espèces mise en place au sein d’un écosystème par une longue coévolution. En effet, comme nous l’avons vu, les changements globaux altèrent les relations entre espèces et modifient la composition du cortège d’espèces présentes au sein des écosystèmes. Le second mécanisme qui risque potentiellement d’être altéré est la complémentarité locale des espèces entre elles (Tilman et al. 2001). En augmentant la diversité, on augmente le nombre de fonctions réalisées par les espèces. De plus, à une échelle régionale, la singularité de chaque communauté entraine une variabilité des réponses aux perturbations qui stabilise globalement tout le système (Olden 2006). Qu’advient-il alors, si les espèces spécialisées disparaissent de l’écosystème et que les mêmes généralistes se retrouvent dans chaque communauté biotique ?
Si nous prenons le recul de l’échelle des temps géologiques, le déclin des espèces spécialistes tend à se retrouver en temps de crise majeure de biodiversité. En effet, les bases de données fossiles montrent que durant les événements d’extinction de masse qu’a connu l’histoire de la vie sur Terre, les espèces spécialistes sont plus fortement touchées et présentent donc une durée de vie sur terre inférieure à des espèces plus généralistes. Le remplacement des spécialistes par des généralistes serait-il alors un stade transitoire avant de retrouver un nouvel assemblage d’espèces nouvellement spécialisées ? Si comme le propose Hutchinson, la biologie de la conservation s’attache à « conserver les acteurs de la pièce évolutive et le décor écologique où elle est jouée », souhaitons alors la bienvenue à ces espèces généralistes.
———-
Bibliographie :
Clavel J., Julliard R. and V. Devictor, 2011. Worldwide decline of specialist species : toward a global functional homogenization? Front. Ecol. Environ. 2 : 222-228.
Devictor V., R. Julliard, J. Clavel, F. Jiguet, A. Lee and D. Couvet. 2008. Functional biotic homogenization of bird communities in disturbed landscapes. Global Ecol Biogeogr. 17: 252-261.
Fisher DO and Owens IPF. 2004. The comparative method in conservation biology. Trends Ecol Evol 19: 391–98
Futuyma DJ and Moreno G. 1988. The evolution of ecological specialization. Annu Rev Ecol Syst 19: 207–33.
Hutchinson GE. 1957. Concluding remarks. CoM Spring Harbor Symp Quant Biol 22: 415–27.
Jablonski D. 2004. Extinction: past and present. Nature 427: 589.
Julliard R, Jiguet F, and Couvet D. 2004. Evidence for the impact of global warming on the long-term population dynamics of common birds. Proc R Soc Lond 271: S490–92.
Krebs C.J.., 2001. Ecology, 5th Edition, Benjamin Cummings.
Olden JD, Poff NL, Douglas MR, et al. 2004. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol Evol 19: 18–24.
Olden JD. 2006. Biotic homogenization: a new research agenda for conservation biogeography. J Biogeogr 33: 2027–39.
Tilman D, Reich P, Knops J, et al. 2001. Diversity and productivity in a long-term grassland experiment. Science 294: 843–45.
Warren MS, Hill JK, Thomas JA, et al. 2001. Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 414 : 65–69
Pour en savoir plus (en français) :
Barbault R. et A. Teyssèdre, 2009. ‘La victime était le coupable’ (Invasions biologiques et bouleversement des écosystèmes). Dossier Pour La Science n°65, octobre 2009, pp. 56-61.
Teyssèdre A, 2007. Dérive et banalisation de l’avifaune. in Quelle Nature voulons-nous ? Observatoires et conservation de la biodiversité, DVDrom, MNHN / distribution CNDP, 2007.
———–
(1) La taille de population est un paramètre important à deux niveaux. Premièrement, d’un point de vue numérique le nombre d’individus introduits comme le nombre d’événements d’introduction sont déterminants dans le succès des espèces exogènes. Deuxièmement, la diversité génétique des populations introduites, qui découle à la fois de la taille de la population initiale et de l’échantillonnage qui en a été fait, favorise une nouvelle fois les espèces généralistes lors de leur établissement.
(2) L’homogénéisation biotique génétique résulte principalement du phénomène d’hybridation entre sous espèces, ou espèces très proches. Les activités humaines responsables de cette HB sont le transport humain via les introductions d’espèces, qui a comme effet d’abattre toutes frontières biogéographiques, et la globalisation de l’agriculture qui au-delà de cultiver les mêmes variétés partout dans le monde au détriment des variétés locales, créer artificiellement par génie génétique des variétés d’organismes génétiquement modifiées qui s’hybrident avec ces voisines sauvages des prés au gré du vent lorsque ces dernières sont de proches cousines.
———-
Ce ‘regard’ est issu de Clavel et al., 2011, “Worldwide decline of specialist species: toward a global functional homogenization?”. In press in Frontiers in Ecology and the Environment.
Regard édité et mis en ligne par Anne Teyssèdre
La conclusion de ce regard est très intéressante et les idées sous-jacentes mériteraient d’être plus diffusées et vulgarisées. Beaucoup d’énergie est dépensée pour la lutte contre les invasives ou la protection des espèces alors que c’est bien la destruction des habitats qui est la plus grande menace en détruisant la « scène » et les « acteurs » des phénomènes écologiques. Est-ce que l’expansion géographique d’espèces généralistes favorisera les diversifications à venir ? C’est une hypothèse intéressante et il serait intéressant de savoir ce qu’en pensent les paléontologues. Les travaux récents sur la diversification récente du genre Cistus (taxon généraliste en méditerranée ?) peuvent peut être fournir quelques pistes (Guzmán, B., Lledó, M. D., and Vargas, P. (2009) Adaptive radiation in Mediterranean Cistus (Cistaceae). PlosOne 4: e6362).
En revanche, sur le début du texte, je ne suis pas sûr que la plus grande vulnérabilité des spécialistes soit généralisable. Est ce qu’il y a eu une synthèse pour les végétaux ?
En 1997, Fisher and Stöcklin ont publié une étude, menée entre 1950 et 1985, portant sur l’extinction locale de différentes espèces de plantes inféodées aux prairies calcaires du Jura Suisse (Fischer and Stocklin 1997). En Europe, les changements de pratiques des sols ont provoqué un déclin drastique des prairies calcaires. De nombreuses espèces de plantes ne persistent plus qu’à l’état de reliques. Sur 35 ans, ils ont trouvé que 39% des populations existantes sur leur site avaient disparu et que les populations les plus exposées à l’extinction locale étaient celles d’espèces spécialistes aux prairies calcaires. De plus, au sein des communautés, ils ont observé un remplacement d’espèces spécialistes par des espèces généralistes. Rooney & al (2004) ont obtenu des résultats similaires dans leur étude portant sur 62 forêts non-fragmentées surveillées pendant plus de 50 ans (Rooney et al. 2004). Ils estiment que ce phénomène de remplacement de spécialistes par des généralistes a augmenté la similarité entre habitats (en termes de composition d’espèces) à hauteur de 9% en moyenne sur ces 50 années.
Pour de la littérature plus récente je vous conseille de lire Fried et al. 2010, BMC Ecology 10:20