La Société Française d’Ecologie (SFE) vous propose cette semaine le regard de Sonia Kéfi, chercheuse à l’Institut des Sciences de l’Evolution de Montpellier (ISEM), sur la dynamique des écosystèmes sous pression et les transitions catastrophiques.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
Des écosystèmes sur le fil :
Comment certains écosystèmes basculent d’un état à un autre
Sonia Kéfi, Institut des Sciences de l’Evolution (ISEM)
Regard R37, édité par Anne Teyssèdre
( Fichier PDF )
Mots clés : transitions catastrophiques, écosystèmes, fonctionnement, résilience, pressions anthropiques, hystéresis, désertification, eutrophisation, boucle de rétroaction positive, point critique, indicateur de dégradation
———
Il reste peu d’endroits sur terre où l’impact de l’Homme n’est pas visible. Presque tous les écosystèmes ont subi ou subissent aujourd’hui des transformations importantes dues aux activités humaines. Ces transformations se sont accélérées au cours des 150 dernières années, durant lesquelles les croissances démographique et économique ont conduit à l’intensification des pressions humaines exercées sur les écosystèmes, telles que l’agriculture, la déforestation et la pêche (voir par ex. les regards n°21 et 30 sur cette plateforme). De plus, en réponse à l’augmentation des concentrations atmosphériques en CO2 et autres gaz à effet de serre, le climat change, exerçant à son tour une pression croissante sur l’ensemble des écosystèmes de la planète 1 (voir par exemple les regards n°10, 22 et 30 sur cette plateforme).
Les activités humaines sont clairement tributaires des écosystèmes, non seulement pour l’approvisionnement en nourriture, mais aussi pour la régulation du climat et des maladies, le contrôle de l’érosion du sol, et la purification de l’eau et de l’air, pour ne donner que quelques exemples. Comprendre comment l’augmentation des pressions exercées sur les écosystèmes affectent – et vont affecter – leur fonctionnement est donc un des enjeux majeurs de l’écologie.
Réponses des écosystèmes aux changements graduels
Lorsqu’une condition environnementale – telle que la quantité de précipitations ou la température atmosphérique – change de façon graduelle au cours du temps (Fig. 1A), on pourrait s’attendre à ce que les écosystèmes répondent eux aussi de façon graduelle (Fig. 1B). Ce n’est pourtant pas toujours le cas. Sous l’effet de pressions croissantes, certains écosystèmes atteignent des points de non-retour au niveau desquels ils subissent des transformations brusques et inattendues appelées « transitions catastrophiques » (ou « catastrophic shifts » en anglais; Fig. 1C) 2.

Figure 1 : Types de réponses d’un écosystème à un changement graduel de condition environnementale. A : Imaginons une condition environnementale qui varie graduellement dans le temps (e.g. quantité de précipitations, température ou apport en nutriments). B – D : Trois types de réponses d’un écosystème à ces changements. L’état de l’écosystème peut correspondre au nombre d’espèces ou à la surface de la couverture végétale par exemple. (B) Transition continue, graduelle : l’état de l’écosystème varie graduellement en réponse au changement de condition environnementale. (C) Transition continue, abrupte : la réponse de l’écosystème devient abrupte et donc moins prévisible mais demeure réversible. (D) Transition discontinue (ou transition catastrophique): l’état du système varie peu jusqu’à ce qu’une valeur seuil de la condition environnementale soit atteinte. L’écosystème bascule alors vers un autre état et donc un autre mode de fonctionnement (par exemple d’un état clair à turbide pour un lac, ou d’un état vert à désertique pour un écosystème aride).
L’eutrophisation des lacs peu profonds est un exemple classique de ce genre de phénomène. Sous l’effet d’une augmentation de l’apport extérieur en nutriments, ces lacs peuvent soudainement passer d’un état riche en végétation submergée et en poissons où l’eau est claire et transparente, à un état eutrophisé où l’eau est trouble, la diversité des espèces bien plus faible, et de moindre attractivité pour le tourisme 3.

Figure 2 : Ecosystème semi-aride : Réserve ornithologique d’El Planerón, Saragosse, Espagne. Les deux photos ont été prises à une centaine de mètres l’une de l’autre. La zone photographiée à droite a été surpâturée jusque dans les années 1950. Ce surpâturage a conduit à une diminution de la couverture végétale, accompagnée d’une érosion du sol, d’une augmentation de la teneur en sel dans le sol et d’un changement de la composition en espèces (dont l’augmentation en fréquence de l’espèce Suaeda vera, tolérante au sel). Suite à sa dégradation, la zone a été abandonnée. Aucune régénération de l’écosystème n’a été observée jusqu’à présent.
Un autre exemple connu de transition catastrophique est la désertification des écosystèmes arides qui peuvent soudainement perdre leur couvert végétal, privant ainsi les populations locales de sources majeures de revenus telles que l’agriculture et le pâturage 4,5 (Fig. 2). Une combinaison de facteurs tels que l’acidification et l’augmentation de la température des océans, la pollution, le tourisme et la surpêche a conduit à la dégradation de certains récifs coralliens 2. Des transitions catas- trophiques ont aussi été observées dans d’autres disciplines telles que l’épidémiologie, la climatologie ou encore l’économie 2.
En provoquant des pertes importantes d’espèces et des réarrangements de communautés écologiques, en affectant le cycle de la matière et de façon plus générale le fonctionnement des écosystèmes, les transitions catastrophiques des écosystèmes peuvent avoir des conséquences écologiques et économiques dramatiques. Les recherches de ces dernières décennies ont donc tenté d’une part de comprendre les mécanismes écologiques qui peuvent expliquer ces comportements et d’autre part d’identifier des signes particuliers annonçant qu’un écosystème se rapproche d’une transition catastrophique. De tels signes pourraient servir à anticiper, voire prévenir, ces transitions.
Les transitions catastrophiques
Une transition catastrophique se produit lorsqu’un écosystème change abruptement d’état (évalué par des variables descriptives, telles que la couverture végétale ou le nombre d’espèces) en réponse à un changement de condition environnementale. Plus précisément, certains écosystèmes restent inertes face à une augmentation de pression jusqu’à ce qu’une valeur seuil, aussi appelée « point critique », soit atteinte à partir de laquelle l’écosystème bascule de son état actuel à un autre (Fig. 1D). La valeur seuil de la condition environnementale étant souvent inconnue, ces transitions se produisent par surprise et sont donc difficiles à anticiper.

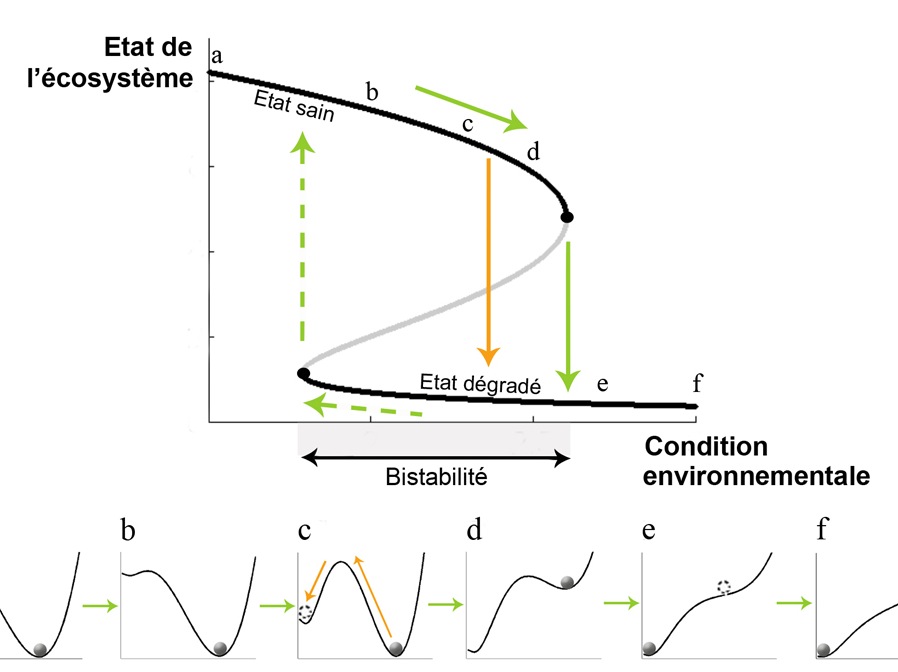
Figure 3. Explication intuitive d’une transition catastrophique. A. Mathématiquement, ce phénomène peut être décrit et expliqué avec des modèles simples (e.g. réf. 6). On parle de bifurcation “fold” ou “saddle-node”, ou encore de transition sous-critique en mathématiques. Ce type de transition se produit lorsque deux états stables d’un écosystème (sain et dégradé) coexistent pour une série de valeurs de la condition environnementale. Ces états stables sont séparés par un équilibre instable (ligne grise) qui marque la limite des bassins d’attraction des deux équilibres stables (lignes noires). B. Paysages de stabilité de l’écosystème (ou « potentiels » en physique) à différents points (a-f) le long du gradient de condition environnementale. Il y a deux façons de passer d’un état à l’autre et donc d’effectuer une transition catastrophique : par modification du paysage de stabilité (flèches vertes) ou par perturbation de l’état de l’écosystème (flèches oranges).
Une des caractéristiques des transitions catastrophiques est qu’il est généralement difficile, voire impossible, de revenir à l’état initial de l’écosystème une fois que la transition s’est produite, même si la pression diminue et si les conditions environnementales reviennent à leur niveau précédant la transition. Il faut une amélioration importante des conditions environnementales pour que l’écosystème retrouve son état et donc son mode de fonctionnement d’origine (Fig. 1D et 3A). Dans le cas des lacs peu profonds par exemple, une fois l’état eutrophisé atteint, une diminution de l’apport en nutriments est souvent insuffisante pour retrouver l’état initial du lac. Le retour de l’écosystème à son état d’origine ne se produit qu’à des niveaux de nutriments beaucoup plus faibles que ceux qui ont conduit à l’eutrophisation (Fig 3A). Ce phénomène dit d’« hystéresis » est dû au fait que dans certaines conditions environnementales le système est bistable, c’est-à-dire qu’il peut être stable dans deux états différents, l’un souvent considéré comme “sain” (eau claire) et l’autre comme “dégradé” (eau trouble).
Le phénomène d’hystéresis est une des caractéristiques importantes des transitions catastrophiques car il est à l’origine de l’irréversibilité possible de la transition une fois qu’elle s’est produite : plus le phénomène d’hystéresis est important (ou plus la zone de bistabilité est large Fig. 3A), plus le retour de l’écosystème à son état d’origine après une transition est difficile. Ce phénomène a donc des conséquences notables pour la conservation et la restauration des écosystèmes.
Une façon imagée de comprendre ce genre de comportement est d’imaginer la trajectoire d’une boule dans un paysage vallonné. Dans un paysage ne comportant qu’une vallée, une boule lâchée du sommet d’une colline ne peut que rouler vers le fond de cette unique vallée et s’y stabiliser (Fig. 3B a,b). En revanche, dans un paysage comportant deux vallées, la trajectoire de la boule – et donc la vallée dans laquelle elle va s’immobiliser – dépend de son point de départ dans le paysage (Fig. 3B c). Dans cette représentation imagée, la boule caractérise l’état de l’écosystème à un instant donné, le paysage correspond à l’ensemble des états dans lesquels l’écosystème peut se retrouver, les « vallées » sont les attracteurs ou équilibres stables, et les sommets des collines sont les équilibres instables.
Un changement de condition environnementale peut provoquer un changement de la forme du paysage, qui peut alors passer d’un relief à deux vallées à un relief à une seule vallée (Fig. 3B d, e). Dans ce cas, une transition catastrophique a lieu lorsque la boule dégringole de la première à la seconde vallée suite à la disparition de la première vallée (Fig. 3 d, e). Dans le cas d’un paysage à deux vallées (Fig. 3B c ou d), une autre façon d’induire une transition catastrophique est de pousser la boule suffisamment fort vers l’amont pour que celle-ci franchisse la crête qui sépare les deux vallées (flèche orange Fig. 3B c ). Ceci correspond à un changement brutal de l’écosystème en réponse à une perturbation importante de son état, telle qu’une diminution drastique du couvert végétal. De la même façon que déplacer une boule d’une vallée à l’autre nécessite beaucoup d’énergie, la remise en état (ou restauration) d’un écosystème dégradé est souvent lente et coûteuse, voire impossible.
Les mécanismes sous-jacents: interactions positives et boucles de rétroaction positives
Quels sont les mécanismes écologiques qui peuvent expliquer de tels comportements à l’échelle des écosystèmes ? La réponse des écosystèmes à des changements de pressions – et en particulier l’émergence de transitions catastrophiques – a été étudiée à l’aide de modèles mathématiques (par exemple réfs 2,6). Ces modèles ont révélé que l’existence de boucles de rétroaction positives entre les organismes vivants et leur environnement abiotique était une cause majeure de réponses abruptes à l’échelle de l’écosystème 2.
Reprenons l’exemple des lacs peu profonds souvent riches en végétation submergée 7. Cette végétation contribue à réduire la turbidité de l’eau en stabilisant le substrat et en protégeant le zooplancton de ses prédateurs, ce qui maintient une faible biomasse de phytoplancton. La végétation submergée favorise ainsi ses propres conditions de croissance en créant une boucle de rétroaction positive entre sa présence et la clarté de l’eau. Au contraire, une perte de végétation peut provoquer une augmentation de turbidité qui défavorise la croissance de la végétation en faisant barrage à la lumière et peut même conduire à sa disparition locale. Des changements de conditions environnementales, tels que l’accroissement d’apports en nitrates et autres nutriments dû à l’agriculture ou des rejets industriels, peuvent activer cette dernière boucle de rétroaction en favorisant la croissance de phytoplancton qui augmente la turbidité de l’eau et diminue le taux de croissance de la végétation submergée. Ceci peut produire des changements brusques du lac d’un état limpide et riche en végétation submergée à un état turbide et pauvre en espèces animales.
De façon similaire, dans les écosystèmes arides, en créant une zone ombragée qui réduit l’évaporation de l’eau et en favorisant la rétention et l’accumulation de matière organique, certaines plantes facilitent la germination et la croissance d’autres individus sous leur canopée 8. (Fig. 2). Ces interactions de facilitation sont susceptibles de créer des boucles de rétroaction positives entre l’abondance de la végétation et la disponibilité locale en ressources limitantes, du même type que celles citées ci-dessus.
Les boucles de rétroaction peuvent conduire à des transitions catastrophiques car elles fonctionnent dans les deux sens : dans le cas des écosystèmes arides, plus de végétation signifie plus de ressources locales qui génèrent à leur tour plus de végétation. En revanche, moins de végétation (par exemple suite à un feu ou à du pâturage intensif) peut inverser la tendance et impliquer moins de ressources, ce qui rend l’installation de nouveaux individus dans l’écosystème de plus en plus difficile. Ces boucles de rétroaction positives peuvent donc contribuer à la stabilisation de l’un ou de l’autre des deux états alternatifs stables de l’écosystème, en fonction du sens dans lequel elles sont activées. Un changement de condition environnementale peut inverser le sens de la boucle de rétroaction et ainsi provoquer un basculement de l’écosystème.
Indicateurs de l’approche d’un point critique
Compte tenu des conséquences potentiellement dramatiques des transitions catastrophiques et de leur caractère peu réversible, de nombreux travaux de recherche ont porté sur l’identification d’indicateurs de transition catastrophique pouvant être utilisés comme des signaux d’alarme précoces, dits « catastrophe flags » ou « early-warning signals » en anglais. Un écosystème qui s’approche d’une transition catastrophique présente-t-il des symptômes particuliers ?
L’idée repose sur un phénomène connu depuis longtemps en physique : quand un système s’approche d’un point critique, il subit un « ralentissement critique » (« critical slowing down » en anglais) 9. En d’autres termes, à mesure qu’un système se rapproche d’un point critique, il requiert de plus en plus de temps pour retrouver son état d’équilibre après une perturbation.
Ce phénomène est visible sur le paysage de stabilité d’un écosystème (Fig. 3 et 4). Lorsqu’un écosystème (représenté par une boule Fig. 3B) s’approche d’un point critique, le fond de la vallée dans laquelle il peut se stabiliser devient de plus en plus plat (Fig. 4A, B). En conséquence, lorsque l’écosystème est perturbé (i.e. lorsque la boule est légèrement déplacée dans le paysage), son retour à son état d’équilibre (i.e. au fond de la vallée) est de plus en plus lent (du fait de la moindre pente des versants).
Indicateurs temporels

Figure 4 : Le ralentissement critique du système à l’approche du point critique. A et B représentent le paysage de stabilité de l’écosystème. C et D montrent des exemples de séries temporelles d’une variable caractéristique du système observées à ces deux points le long de la transition catastrophique. La série temporelle D est plus variable, plus autocorrélée et plus asymétrique que la série temporelle C.
Le phénomène de ralentissement critique a des conséquences concrètes quantifiables sur la dynamique du système 10. Tout d’abord, la variabilité du système augmente, car suite à une perturbation dans une vallée au fond plus plat l’amplitude des déplacements de la boule est plus importante – en d’autres termes, l’écosystème explore une plus grande variété d’états 11. D’autre part, les états du système à des instants successifs t et t+1 se ressemblent d’avantage, car la boule se déplace plus lentement 12. Enfin, comme la forme du paysage change et devient plus asymétrique à l’approche d’un point critique (Fig. 3d), la boule va avoir tendance à passer plus de temps d’un côté de la vallée que de l’autre suite à une perturbation13. Le suivi temporel d’une variable caractéristique de l’écosystème, telle que la biomasse ou la couverture végétale, présente alors des fluctuations de plus en plus amples, auto-corrélées et asymétriques à l’approche du point critique, comme l’illustrent les figures 4C et D ci-dessous. La variance, l’autocorrélation et l’asymétrie temporelle de ces variables descriptives peuvent donc être utilisées comme indicateurs temporels de l’approche d’un point critique.
Indicateurs spatiaux
Récemment, des études ont montré que les équivalents spatiaux de ces indicateurs 14-16 se comportent de la même façon que les indicateurs temporels. En d’autres termes la variance spatiale, l’auto-corrélation spatiale et l’asymétrie spatiale de variables caractéristiques d’un écosystème pourraient annoncer l’approche d’un point critique.
D’autre part, dans les écosystèmes qui présentent une structure spatiale claire (par exemple, un couvert végétal fragmenté comme c’est souvent le cas dans les écosystèmes arides ; Fig. 2), des indicateurs spécifiques à ces écosystèmes ont été proposés et pourraient donner une idée du niveau de dégradation de l’écosystème 17-19 (par ex. la taille et la forme des éléments de végétation dans le cas des écosystèmes arides).
Limitations de ces indicateurs
En résumé, lorsque des données temporelles ou spatiales qui suivent une variable caractéristique d’un écosystème sont disponibles, l’évaluation de quantités relativement simples (variance, autocorrélation et asymétrie) peut donner des informations sur l’approche d’un point critique. Des preuves expérimentales de la détectabilité de ces indicateurs, initialement développés à l’aide de modèles mathématiques, ont récemment été publiées 20-23, 27. Ces indicateurs semblent donc prometteurs. Cependant de nombreux problèmes techniques persistent, qui doivent être résolus avant de pouvoir utiliser ces indicateurs comme de véritables outils pour la gestion des écosystèmes.
Tout d’abord, aucun des indicateurs présentés ci-dessus ne permet d’évaluer quantitativement la distance au point critique. Même si ces indicateurs suggèrent que l’écosystème s’approche d’un point critique, ils ne donnent aucune information sur la quantité de perturbation ou de pression externe que l’écosystème peut encore subir avant de basculer. D’autre part, ces indicateurs ont été développés à partir de modèles mathématiques relativement simples et la question de leur détectabilité dans des systèmes complexes demeure 24. Enfin, leur détection statistique dans des écosystèmes réels reste un défi, d’une part parce que les données disponibles n’ont pas toujours une résolution suffisante, d’autre part parce qu’on ne dispose pas encore de routine statistique pour leur application.
De façon plus générale, les modèles mathématiques suggèrent que ce comportement de ralentissement critique se produit à l’approche de différents types de transitions, et ne sont donc pas propres au cas des transitions catastrophiques telles qu’illustrées en Fig. 1D et 3A. Des signaux semblables (i.e. ralentissement, auto-corrélation, amplification et asymétrie des variations) peuvent en effet aussi être détectés dans le cas d’une réponse abrupte mais réversible de l’écosystème à un changement de condition externe, telle qu’illustrée Fig. 1C. Une augmentation de la valeur des indicateurs suggère donc que le système devient de plus en plus fragile face aux changements de pressions qu’il subit, car il peine à retourner à son état d’origine après une perturbation. Par contre, ils n’annoncent pas nécessairement l’approche d’un point critique au niveau duquel l’écosystème peut basculer de façon discontinue et irréversible. En d’autres termes, les indicateurs ne procurent pas d’information quant à la réversibilité de la transition qui approche.
Conclusion
La conférence des Nations Unies 2012 pour le Développement Durable (Rio+20) s’est tenue à Rio au Brésil du 20 au 22 juin dernier. Achim Steiner, directeur général du Programme des Nations Unies pour l’Environnement (PNUE) a souligné à cette occasion que « si cette situation perdure, si les structures actuelles de production et de consommation des ressources naturelles continuent à prévaloir et que rien n’est fait pour inverser la tendance, les gouvernements devront assumer la responsabilité d’un niveau de dégradation et de répercussions sans précédent ».
Si les transitions catastrophiques évoquées dans ce « regard » sont observées à l’échelle locale, des transitions planétaires (dites globales) se sont produites à l’échelle des temps géologiques, par exemple lors de la dernière transition glaciaire-interglaciaire il y a un peu plus de 10000 ans (28 et voir le regard n°10 de Valérie Masson-Delmotte sur cette plateforme). Ces transitions globales ont été provoquées par des forçages à l’échelle planétaire. Les changements que nous imposons aujourd’hui sur la planète – croissance de notre population, transformation et fragmentation des habitats, changements climatiques, consommation des ressources naturelles… – sont-ils de la même envergure ? Dans un article récemment publié dans la revue Nature 28, Barnosky et ses collègues suggèrent qu’ils le sont, voire qu’ils les dépassent à la fois en rapidité et en amplitude.
Si Barnosky et collègues voient juste, la planète pourraient se rapprocher d’un point critique. Une transition globale n’est cependant peut-être pas inévitable. Anticiper la réponse des écosystèmes aux changements globaux et prévenir leur dégradation irréversible font partie des défis des décennies à venir 25. Relever ces défis repose sur les choix politiques faits dans les quelques années à venir aux échelles nationales et internationales, mais aussi sur les avancées de la science, et particulièrement de l’écologie. Les travaux de recherche sur les transitions catastrophiques et les indicateurs de points critiques participent à améliorer notre compréhension de la persistance et de la stabilité des écosystèmes et visent, à terme, à fournir des outils de prédiction, d’anticipation et de gestion permettant à nos sociétés de faire face aux défis environnementaux croissants 26.
Bibliographie
1. Houghton, J.T. et al. Climate change 2001: The scientific basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge (2001).
2. Scheffer, M., Carpenter, S.R., Foley, J.A., Folke, C. & Walker, B. Catastrophic shifts in ecosystems. Nature 413, 591-6 (2001).
3. Carpenter, S.R., Ludwig, D. & Brock, W.A. Management of eutrophication for lakes subject to potentially irreversible change. Ecological Applications 9, 751-771 (1999).
4. Rietkerk, M., Ketner, P., Stroosnijder, L. & Prins, H.H.T. Sahelian rangeland development; a catastrophe ? Journal Of Range Management 49, 512-519 (1996).
5. Reynolds, J.F. et al. Global desertification: Building a science for dryland development. Science 316, 847-851 (2007).
6. May, R.M. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 269, 471-477 (1977).
7. Scheffer, M., Hosper, S.H., Meijer, M.-l & Jeppesen, E. Alternative equilibria in shallow lakes. Trends in ecology & evolution 8, 275-9 (1993).
8. Aguiar, M.R. & Sala, O.E. Patch structure, dynamics and implications for the functioning of arid ecosystems. Trends in Ecology and Evolution 14, 273-277 (1999).
9. van Nes, E.H. & Scheffer, M. Slow recovery from perturbations as a generic indicator of a nearby catastrophic shift. The American naturalist 169, 738-47 (2007).
10. Scheffer, M. et al. Early-warning signals for critical transitions. Nature 461, 53-9 (2009).
11. Carpenter, S.R. & Brock, W.A. Rising variance: a leading indicator of ecological transition. Ecology letters 9, 308-315 (2006).
12. Held, H. & Kleinen, T. Detection of climate system bifurcations by degenerate fingerprinting. Geophysical Research Letters 31, L23207 (2004).
13. Guttal, V. & Jayaprakash, C. Changing skewness: an early warning signal of regime shifts in ecosystems. Ecology letters 11, 450-60 (2008).
14. Dakos, V., Nes, E.H., Donangelo, R., Fort, H. & Scheffer, M. Spatial correlation as leading indicator of catastrophic shifts. Theoretical Ecology 3, 163-174 (2009).
15. Guttal, V. & Jayaprakash, C. Spatial variance and spatial skewness: leading indicators of regime shifts in spatial ecological systems. Theoretical Ecology 2, 3-12 (2008).
16. Dakos, V., Kéfi, S., Rietkerk, M., van Nes, E.H. & Scheffer, M. Slowing down in spatially patterned ecosystems at the brink of collapse. The American naturalist 177, E153-66 (2011).
17. Rietkerk, M., Dekker, S.C., de Ruiter, P.C. & van de Koppel, J. Self-organized patchiness and catastrophic shifts in ecosystems. Science 305, 1926-9 (2004).
18. Kéfi, S. et al. Robust scaling in ecosystems and the meltdown of patch size distributions before extinction. Ecology letters 14, 29-35 (2011).
19. Kéfi, S. et al. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 449, 213-7 (2007).
20. Dakos, V. et al. Slowing down as an early warning signal for abrupt climate change. Proceedings of the National Academy of Sciences of the United States of America 105, 14308-12 (2008).
21. Drake, J.M. & Griffen, B.D. Early warning signals of extinction in deteriorating environments. Nature 467, 456-9 (2010).
22. Carpenter, S.R. et al. Early warnings of regime shifts: a whole-ecosystem experiment. Science 332, 1079-82 (2011).
23. Veraart, A.J. et al. Recovery rates reflect distance to a tipping point in a living system. Nature 481: 357-359 (2011).
24. Hastings, A. & Wysham, D.B. Regime shifts in ecological systems can occur with no warning. Ecology letters 13, 464-72 (2010).
25. Millennium Ecosystem Assessment Ecosystems and Human Well-Being: Desertification Synthesis. World Resources Institute, Washington DC. (2005).
26. Loreau, M. Linking biodiversity and ecosystems: towards a unifying ecological theory. Philosophical transactions of the Royal Society of London. Series B, Biological sciences 365,
49-60 (2010).
27. Dai, L. et al. Generic Indicators for Loss of Resilience Before a Tipping Point Leading to Population Collapse. Science 336 (6085) 1175-1177 (2012).
28. Barnosky, A.D. et al. Approaching a state shift in Earth’s biosphere. Nature 486, 52–58 (2012).
—
Pour en savoir plus, en français :
Ces trois regards sur l’ampleur, l’impact et les enjeux du changement climatique actuel :
Masson-Delmotte V., 2011. Le climat de notre biosphère. Regards et débats sur la biodiversité, SFE, Regard n°10, janvier 2010.
Julliard R. et F. Jiguet, 2011. La biodiversité face au changement climatique : Ce que nous indique les oiseaux. Regards et débats sur la biodiversité, SFE, Regard n°22, septembre 2011.
Lavorel S., R. Barbault et J-C. Hourcade, 2012. Impact et enjeux du changement climatique. Regards et débats sur la biodiversité, SFE, Regard n°30, mai 2012.
Ce chapitre sur la stabilité, la résilience et le basculement des écosystèmes :
Couvet D. et Teyssèdre A., 2010. Ecologie et biodiversité, des populations aux socioécosystèmes, chapitre 5 (pp.162 à 185). Belin, juin 2010.
Article édité par Anne Teyssèdre
——
Bonjour Sonia,
J’ai lu avec beaucoup d’intérêt vos travaux sur les lois de distribution des ‘patchs’ dans certains milieux arides, et la théorisation correspondante pour définir des indicateurs de l’état de dégradation (désertification) de ces milieux. Existe-t-il des travaux, publiés ou en cours, sur l’application pratique de cette démarche pour la cartographie régulière (et le monitoring) de l’état de dégradation? Est il déjà possible d’évaluer concrètement l’aide que cela pourrait apporter dans les actions orientées vers la gestion durable des terres ?
Bonjour,
Merci beaucoup pour vos questions. Il n’y a pas encore de travaux publiés sur l’utilisation pratique des indicateurs spatiaux de transitions catastrophiques. Il s’agit en effet tout d’abord de les valider, et plusieurs équipes y travaillent actuellement. Dans le cas des distributions de tailles de patchs, nous avons besoin de séries temporelles d’images aériennes (i) de résolution suffisante pour détecter ces structures qui sont souvent petites dans les écosystèmes Méditerranéens (de l’ordre du mètre) et (ii) qui couvrent une période de temps au cours de laquelle une dégradation s’est produite (l’idéal serait de trouver une zone considérée comme dégradée aujourd’hui et de pouvoir trouver des images à plusieurs instants dans le temps avant la dégradation). Une telle série temporelle nous permettrait d’observer les changements d’organisation spatiale de la végétation avant la désertification de l’écosystème, de vérifier que les changements de structures observés dans les modèles sont effectivement observés sur le terrain, et de quantifier les échelles de temps sur lesquels ces changements sont observables. De tels jeux de données diachroniques (à plusieurs pas dans le temps) de qualité suffisante pour pouvoir réaliser ces analyses sont rares.
Il y a par ailleurs des travaux très intéressants sur les structures régulières de végétation, pour lesquelles – parce que beaucoup plus larges – il est plus facile de trouver des jeux de données. Dans ce cas, la forme des patchs de végétation change le long d’un gradient de stress (climat ou impact humain) (Rietkerk et al. 2004) : le système passe d’un couvert végétal continu à des « gaps » (couvert végétal troué), des labyrinthes puis des « spots ». Barbier et collègues (2006) ont effectivement observés la transition d’un couvert végétal homogène à un couvert troué (« gaps ») suite à une diminution de pluviométrie dans le Sahel entre 1956 et 1996. Deblauwe et collègues (2011) ont observé les changements de gaps, à labyrinthes, et à spots le long d’un gradient de pluviométrie dans l’espace et dans le temps au Soudan. Ces études valident les prédictions des modèles théoriques et confirment que les spots sont les dernières structures observées avant la désertification des écosystèmes caractérisés par une organisation régulière de la végétation.
Sonia
Références :
Barbier, N. et al. Self-organized vegetation patterning as a fingerprint of climate and human impact on semi-arid ecosystems. Journal of ecology 94(3), 537-547 (2006)
Deblauwe, V. et al. The global biogeography of semi-arid periodic vegetation patterns. Global Ecology and Biogeography 17(6), 715-723 (2008)
Deblauwe, V. et al. Environmental modulation of self-organized periodic vegetation patterns in Sudan. Ecography 34(6): 990–1001 (2011)
Rietkerk, M. et al. Self-Organized Patchiness and Catastrophic Shifts in Ecosystems. Science 305: 1926-1929 (2004)
Bonjour,
Pour commencer, je tiens à vous remercier pour cet article très clair et pédagogique (compréhensible par des profanes).
Je me permets de vous poser quelques petites questions relatives à la place de l’homme quant aux basculements des écosystèmes.
Concernant les écosystèmes qui connaissent des changements graduels (j’écarte le cas des transitions catastrophiques qui sont radicales et irréversibles) : n’est-il pas possible que le caractère progressif du changement permette aux espèces de l’écosystème d’évoluer et de s’adapter naturellement (je suppose que le temps est un facteur important)? En effet, il me semble que l’évolution est aussi une forme de sélection des espèces végétales et animales les plus résistantes. Il me semble que dans un premier temps de nombreuses espèces vont disparaitre et seules les plus résistantes pourront survivre. Mais n’y aura-t-il pas un second temps où les espèces résistantes vont évoluer et se diversifier (je suppose que le facteur temps est là aussi très important)?
Enfin, l’homme qui aujourd’hui a de plus en plus les moyens de détecter les « catastrophe flags » ne peut-il pas agir sur les différents éléments des boucles de rétroaction positives afin de favoriser les basculements des écosystèmes (pas spécialement vers ce que la nature avait mis en place)? Et l’intervention de l’homme dans ces changements n’est-elle pas le dernier des aménagements de l’habitat de l’homme?
Bien cordialement
Bonjour,
L’évolution des espèces est effectivement un maillon manquant de la théorie des transitions critiques. Si les variations de conditions environnementales sont suffisamment lentes pour que les espèces aient le temps de s’y adapter, il pourrait y avoir des phénomènes de « sauvetage évolutif ». Tout dépend des échelles de temps relative du changement environnemental et de l’adaptation.
Concernant votre seconde question, les boucles de rétroaction positives sont effectivement utilisées en écologie de la restauration pour faciliter le retour d’un écosystème qui a subit une transition catastrophique à son état initial. Suding et collègues ont écrit une excellente revue à ce sujet (Suding et al. 2004).
Sonia
Référence: Suding et al. 2004. Alternative states and positive feedbacks in restoration ecology. Trends in ecology and evolution 19(1): 46-53
Bonjour
et merci pour ce regard qui finalement explique très bien des choses théoriques compliquées. J’ai une question qui rejoint un peu les deux autres commentaires. Il y a t il des cas concrets où des indicateurs ont pu être utilisés pour prédire voir prévenir une transition catastrophique?
Cordialement,
Sébastien
Bonjour,
Les premières preuves expérimentales des indicateurs de transitions catastrophiques sont récentes. En laboratoire, Drake et Griffen (2010) ont conduit des populations de Daphnées à l’extinction et ont observé une augmentation de la variance, de l’asymétrie (skewness) et de l’autocorrélation (temporelle) avant le point critique. Veraart et al. (2011) ont montré le ralentissement critique de populations de cyanobactéries en route vers l’extinction, associé à une augmentation de l’autocorrélation temporelle. Dai et al. (2012) ont observé un ralentissement critique de la dynamique de levures à l’approche d’un point critique.
A plus large échelle, Steve Carpenter et ses collègues ont expérimentalement fait basculer tout un réseau trophique dans un lac. Ils ont montré le ralentissement critique des populations, ainsi qu’une augmentation de la variance, de l’autocorrélation, et de la skewness avant le point critique.
Enfin, ces indicateurs semblent être détectables dans des données de séries temporelles de climat avant les grands changements climatiques passés (e.g. Dakos et al. 2008) .
Sonia
Références:
Carpenter et al. Early warnings of regime shifts: a whole-ecosystem experiment. Science 332(6033): 1079-1082 (2011)
Dai et al. Generic indicators for loss of resilience before a tipping point leading to population collapse. Science 336: 1175-1177 (2012)
Dakos et al. Slowing down as an early warning signal for abrupt climate change. PNAS 105: 14308 (2008)
Drake and Griffen. Early warning signals of extinction in deteriorating environments. Nature 467: 456 (2010)
Veraart et al. Recovery rates reflect distance to a tipping point in a living system. Nature 481: 357 (2011)
Bonjour à tous,
Cet article est important en ce qu’il rappelle que les écosystèmes ne sont pas statiques. Nous avons tous encore trop tendance à les étudier fixes, avec l’illusion qu’ils sont dans un état d’équilibre ou pire, dans un instantané figé. Toutefois, on peut pousser plus loin la logique de ce document.
Tant que l’on parvient à formaliser les dynamiques écosystémiques par des équations différentielles, cela signifie que l’on dispose d’un même paramètre à partir duquel on peut suivre quantitativement l’évolution de l’écosystème, en particulier de sa composition et sa structure. Dans ce cas, les processus écologiques à l’œuvre sont aujourd’hui bien connus (colonisation, variations biogéochimiques…) et peuvent soutenir ces modèles plutôt phénoménologiques de ruptures violentes.
Mais on peut envisager des changements plus catastrophiques encore, ceux qui bousculeront assez l’écosystème pour en changer la structure, en un point par exemple où la résolution des équations de sa dynamique ne sera plus possible. C’est certainement ce qu’il se passe lorsque les écosystèmes changent de nature, (on tente de capturer ces changements avec un indicateur d’ouverture du milieu ou de biomasse, mais qui n’a plus d’interprétation écologique directe) : c’est le cas d’espèces invasives virulentes et de disparitions en chaine qui transforment le réseau trophique, ou encore de réduction de la surface de l’écosystème (e.g. par érosion ou clôture d’une réserve par l’homme). Dans ces cas, c’est la structure même des interactions entre les composantes écosystémiques qui est rompue.
Nous avons sûrement encore trop peu de données pour quantifier de telles ruptures, mais il y a fort à parier que de tels bouleversements sont déjà légions avec l’emprise de l’homme sur son environnement, comme ce document le rappelle bien. Or, on ne sait pas anticiper (prédire) ces catastrophes. Nous n’avons pas les outils mathématiques et conceptuels pour nous y attaquer ! Comment par exemple résoudre des équations sur une structure changeante ? D’autres disciplines se sont interrogées. C’est probablement à cette question théorique que l’écologie des écosystème doit s’atteler !
Cédric
Bonjour,
Merci pour ce regard très intéressant. Je me posais une question sur les indicateurs de changements d’état.
On découvre des exemples de détection à partir d’indicateurs qui peuvent porter à des échelles différentes: écosystémiques (ex: structure de végétation) ou bien à l’échelle de populations d’espèces (ex: populations de daphnées ou de cyanobactéries). Je me demandais si une échelle d’organisation (population, espèce, écosystème) semblait plus « sensible » que d’autres pour la détection de points critiques dans le fonctionnement des écosystèmes? Y compris d’un point de vue théorique?
Dans la pratique, on commence à disposer de plus en plus de données standardisées sur le suivi d’abondances d’espèces ou bien de l’état de conservation de certains écosystèmes à des échelles « locales ». On pourrait imaginer qu’elles puissent être utilisées à terme par les gestionnaires d’espaces naturels pour appréhender ces changements d’état.
Merci,
Baptiste
Bonjour,
C’est une question très importante. A ma connaissance, les études théoriques se sont peu penchées sur la question, même si, de fait, différentes études théoriques quantifient les indicateurs sur des variables correspondant à différents niveaux d’organisation: les études sur la désertification des écosystèmes arides suivent souvent la biomasse totale de végétation ou le couvert végétal, i.e. des caractéristiques évaluées à l’échelle de l’écosystème (e.g. Kéfi et al. 2007; Guttal et Jayaprakash 2008); d’autres études suivent les abondances de population (e.g. Drake et al. 2010; Veraart et al. 2012).
Le choix de ces variables caractéristiques dépend du type de transition observée ; en fonction des caractéristiques du système étudié, une variable s’impose souvent plus qu’une autre. Cependant, ce choix repose, au moins en partie, sur l’intuition des chercheurs spécialistes du système. Des études théoriques qui aborderaient ces questions de façon systématique seraient extrêmement utiles.
Sonia
Bonjour,
Merci pour cet article intéressant. Je m’interroge sur la présentation somme toute assez positive que vous faites des indicateurs temporels et/ou spatiaux pour les changements de régimes, dans l’article et les commentaires.
La majorité des « validations » de ces indicateurs proviennent en effet de simulations sur des variantes du modèle de May (1977) (modèle logistique avec récolte, fonction d’extraction de forme GX^p/(X^p+D^p)), sur des séries temporelles presentant des changements, et cela pour plusieurs centaines de points. Indépendamment des problèmes liés à la spécificité du modèle, le processus de collecte des données empiriques est souvent insuffisant, comme vous le mentionnez, pour utiliser ces indicateurs. Un collègue m’a fait suivre ce papier – qu’en pensez-vous?
Perretti, Charles T., and Stephan B. Munch. 2012. Regime shift indicators fail under noise levels commonly observed in ecological systems. Ecological Applications 22:1772–1779. http://dx.doi.org/10.1890/11-0161.1
A l’inverse, sur des données riches, un article récent signale une quantité importante de faux positifs:
Boettiger and Hastings (2012). Early Warning Signals and the Prosecutor’s Fallacy. Proceedings of the Royal Society B (in press). doi:10.1098/rspb.2012.2085
La conclusion des études par simulation semble donc que même sur le modèle de May et ses dérivés, il n’est pas vraiment possible d’identifier en pratique des transitions critiques, en raison de l’échantillonnage et de la stochasticité inhérente aux systèmes écologiques. De plus, comme indiqué dans le papier de Hastings et Wysham (et le votre dans Oikos?), beaucoup de systèmes n’ont pas de bifurcation « saddle-node » comme le modèle de May, et/ou ont des bifurcations complétement différentes…
Travaillant actuellement sur des populations cycliques de micro-mammifères pour lesquelles la variance en abondance change quasi perpétuellement (et avec elle, celles de bien d’autres espèces dans l’écosystème entier), je trouve le critère d’augmentation de la variance particulièrement pernicieux. En ce qui concerne les indicateurs spatiaux, bon nombre de modèles spatiaux (e.g. prédateurs-proies, avec ou sans cycles limites) peuvent montrer des changements de covariance spatiale marqués sans pour autant se diriger vers une transition critique ni un profond changement d’etat. Bref, il me semble tres difficile d’utiliser les critières EWS pour un grand nombre d’organismes / ecosystemes.
Etant donné que Kendall et al. 1998 ont estimé vers 30% la proportion de séries temporelles cycliques dans la Global Population Dynamics Database, 30% semble une minoration de l’ensemble des séries temporelles sur lesquelles chercher des early-warning signals n’a pas vraiment de sens écologique.
Quelles restrictions suggèreriez-vous sur l’usage de ces indicateurs? Ce genre d’information pourrait sans doute être utile aux chercheurs utilisant des données empiriques.
Cordialement,
Frédéric
Bonjour,
Effectivement, les indicateurs (EWS) ont initialement été développés sur des modèles mathématiques très simples, si bien que la détectabilité de ces indicateurs dans des systèmes réels se pose. Le fait est que ces indicateurs ont été détectés dans plusieurs systèmes réels, complexes (e.g. Carpenter et al. 2011). Cependant, on peut imaginer qu’il y ait un certain nombre de cas dans lesquels les signaux ne se produiront pas ou ne seront pas détectables. Quels sont ces cas? Dans quels systèmes a-t-on une chance de détecter ces indicateurs? Plus de travail théorique est nécessaire pour élucider ces questions. Récemment certaines études ont émis des doutes quant à la détectabilité des indicateurs dans des systèmes complexes (Hastings 2009). L’article que vous citez, Perreti et Munch 2012, montre que les signaux des indicateurs EWS sont faibles dans des systèmes avec beaucoup de bruit.
Dans un article qui va bientôt paraitre dans Oikos, nous avons étudié le comportement des indicateurs EWS quand un système (le modèle de May 1977) se rapproche de différents types de points critiques dont un seul correspond à une transition catastrophique. Nous montrons que les indicateurs sont positifs dans tous ces cas. En d’autres termes, ils ne sont pas spécifiques aux transitions catastrophiques mais semblent indiquer de façon plus générale le rapprochement d’un point critique, quelle que soit sa nature.
Cela conduit à nous poser la question suivante: si un signal est détecté, qu’est-ce que cela signifie? Tout d’abord, les études théoriques suggèrent qu’on peut plus se fier à une combinaison d’indicateurs qu’à des indicateurs isolés. De nombreux phénomènes peuvent en effet être à l’origine d’une augmentation de variance dans une population par exemple. Cependant si des augmentations de variance, d’auto-corrélation et d’asymétrie sont simultanément observées, les chances sont importantes que cela signale un ralentissement critique du système, et donc une diminution de la résilience du système.
Le développement de ces indicateurs est encore à un stade de recherche fondamentale. Des critères clairs pour leur application dans des cas réels ne sont donc malheureusement pas encore disponibles mais en cours de développement. Des études théoriques sont nécessaires pour identifier les conditions dans lesquelles ils peuvent être observés, en parallèle avec des validation sur des données.
Je me permets de vous signaler la parution d’un article récent sur le sujet publié par Boettinger et Hastings dans Nature:
http://www.nature.com/nature/journal/v493/n7431/full/493157a.html
Merci pour cet article passionnant.
Je m’intéresse (à titre personnel) à la question de la modification des écosystèmes forestiers, notamment en milieu tempéré. Savez-vous s’il existe des travaux de recherche sur ce sujet, en particulier concernant l’instrumentation en vue d’une transition catastrophique dans cet écosystème ? Pensez-vous qu’une telle transition pourrait arriver sur les forêts en milieu tempéré, et quels en seraient les indicateurs ?
Merci beaucoup pour vos précisions.
Bonjour,
Je ne connais pas d’étude sur les transitions catastrophiques en écosystèmes forestiers de milieu tempéré.
En 2011, Hirota et collègues ont publié un article sur les forêts tropicales et savanes: http://www.sciencemag.org/content/334/6053/232.short
Plus récemment, Marten Scheffer et collègues ont publié un article sur les forêts boréales que vous trouverez peut-être intéressant: http://www.pnas.org/content/early/2012/12/06/1219844110.abstract