La Société Française d’Ecologie (SFE) vous propose cette semaine le regard de Christelle Jozet-Alves, chercheuse en éthologie à l’Université de Caen, sur le sens du temps – mémoire épisodique et planification – chez les animaux. Ce regard est une version légèrement modifiée et augmentée d’un article de la même auteure, paru dans le n°2 de la revue ESpèces, partenaire de ce projet.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires après cet article; les auteurs vous répondront.
Les animaux se projettent-ils dans le temps ?
Christelle Jozet-Alves
Groupe mémoire et Plasticité comportementale,
Université de Caen Basse-Normandie
Regard R29, édité par Anne Teyssèdre
———
Mots clés : mémoire épisodique, anticipation, temps, comportement animal,
évolution, biodiversité
———
Est-ce que les animaux se souviennent du bon vieux temps et méditent sur ce que leur réserve l’avenir ? Cette question à première vue ésotérique est le point central de débats animés au cœur de la communauté scientifique. L’Homme est capable de se remémorer des évènements passés, et de s’imaginer ce que pourrait être son futur. La mémoire des évènements vécus est appelée « mémoire épisodique » (par exemple : je me souviens de mon dernier voyage à Londres), elle se distingue de la « mémoire sémantique » qui permet le stockage de la connaissance générale sur le monde (par exemple : je sais que Londres est une capitale européenne).
Revivre des évènements passés et se projeter dans le futur sont des « voyages mentaux dans le temps » à partir du moment présent. Ces voyages mentaux contrôlent notre comportement quotidien : « je prévois d’acheter du beurre demain, car je me souviens avoir remarqué, ce matin, qu’il n’en restait plus beaucoup ». Suddendorf et Corballis (1997) ont suscité une importante polémique en affirmant que cette capacité serait une caractéristique unique de l’espèce humaine. D’après ces auteurs, les animaux seraient « ancrés » dans l’instant présent, ils n’auraient pas la notion du temps. Ceci suggèrerait une discontinuité très nette dans l’évolution des capacités cognitives au sein du règne animal. En réaction, ces affirmations ont généré un nombre important d’articles plus ou moins controversés et ont donné une impulsion forte à de nouvelles pistes de recherche chez l’animal.
La mémoire épisodique chez les animaux
Il est impossible d’interroger les animaux pour savoir s’ils se remémorent consciemment un évènement (mémoire épisodique) ou s’ils « savent » ce qu’ils ont vécus (mémoire sémantique, pas de voyage mental dans le temps ; voir le dessin ci-dessous de Ruth Tulving). Ainsi, en 1998, Clayton et Dickinson ont préféré employer l’expression mémoire « de type » épisodique, pour parler de la mémoire des évènements chez l’animal. Pour la mettre en évidence, des critères purement comportementaux ont été utilisés, notamment l’existence de trois composantes : le « quand », le « quoi » et le « où » dans la mémoire d’un événement.

Dessin de Ruth Tulving
Ce dessin de Ruth Tulving résume la problématique engendrée par l’étude de la mémoire épisodique chez les animaux, à savoir l’impossibilité de les interroger pour savoir s’ils « savent » ce qu’ils ont vécu ou s’ils se remémorent consciemment un évènement.
Il semble très avantageux pour un prédateur de se souvenir a) de la localisation des sources de nourriture qu’il a rencontrées par le passé, b) s’il restait des proies après son passage, et c) combien de temps s’est écoulé depuis. En effet, si toutes les proies ont été consommées à un en-droit donné, l’animal pourra éviter d’y retourner dans les jours qui suivent, et y reviendra après un certain délai puisqu’il se souviendra qu’il y a souvent des proies cachées à cet endroit.
Clayton et ses collaborateurs ont engagé une série d’expériences très innovantes chez l’oiseau. Ils se sont appuyés sur le comportement naturel de geais qui stockent leur nourriture pour tester leur capacité à se souvenir quel type de nourriture, ils ont stocké « où » et depuis combien de temps (Clayton & Dickinson 1998, 1999 ; Clayton et al. 2001). Dans leurs paradigmes expérimentaux, des geais à gorge blanche (Aphelocoma coerulescens) apprennent à resituer dans le temps et l’espace plusieurs types d’items alimentaires : certains très appréciés mais périssables (comme les vers de terre par exemple), d’autres moins appréciés mais plus durables (exemple : cacahuètes). En fonction du temps écoulé depuis qu’ils ont caché de la nourriture, les geais ajustent de manière très appropriée leur recherche d’aliments, démontrant ainsi qu’ils prennent en compte l’écoulement du temps dans leurs comportements alimentaires (voir la Figure 1 ci-dessous).
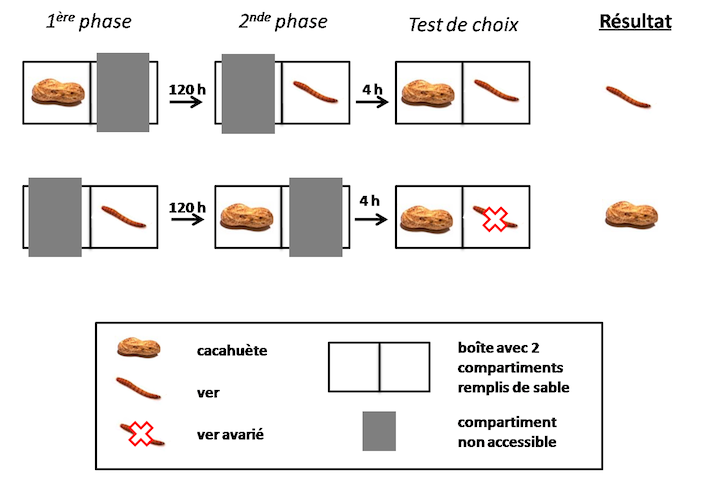
Figure 1 : Procédure expérimentale utilisée par Clayton et Dickinson (1998). Les geais préfèrent les vers, mais ils ont appris au préalable que les vers se détériorent avec le temps. 1ère phase : les geais sont en présence soit de cacahuètes, soit de vers qu’ils peuvent enfouir dans le sable d’un des deux compartiments d’une boîte. Après 120 h, 2nde phase : le deuxième type d’item alimentaire leur est proposé, les geais peuvent le flushr dans le deuxième compartiment. 4 h plus tard, les geais sont placés devant la boîte où les deux compartiments sont accessibles. Les auteurs ont montré que les oiseaux cherchent de la nourriture en fouillant le sable du côté « vers » lorsque ceux-ci ont été cachés depuis peu de temps, et du côté « cacahuète » lorsque les vers ont été cachés depuis longtemps (124 h).
La capacité de se souvenir des évènements passés pourrait également être mise à profit au cours des interactions sociales. Chez les espèces sociales, des relations hiérarchiques se mettent en place en fonction des interactions entre les individus. Ainsi, pour exprimer un comportement social approprié, un animal doit se souvenir des évènements qui auraient modifié les relations hiérarchiques pré-existantes (qu’est-ce qu’il s’est passé, quand, où et qui était impliqué). Ferkin et ses collaborateurs (2008) ont montré que chez le campagnol des champs, une espèce polygyne (i.e., chez laquelle les mâles reproducteurs peuvent s’accoupler avec plusieurs femelles), les mâles connaissent la localisation de différentes femelles et parviennent à caler leurs visites sur la période de réceptivité sexuelle de chacune de ces femelles.
Ces études suggèrent que des animaux peuvent voyager mentalement dans le passé dans un cadre temporel de plusieurs heures, voire de plusieurs jours. Certains auteurs exposent toutefois une hypothèse alternative (Roberts et al. 2008) : les animaux « sauraient » ce qu’il s’est passé (mémoire sémantique) et garderaient simplement en mémoire le temps écoulé en se référant, par exemple, à l’intensité du souvenir qui diminuerait au cours du temps. En fonction de cette durée, les animaux retourneraient ou non près de la source de nourriture ou près de leurs congénères. Ainsi, pour ces auteurs, il ne s’agirait pas d’une reconstruction mentale d’évènements vécus, comme ce qui est observé chez l’Homme.
Se souvenir du bon temps…
Le composant temporel de la mémoire de type épisodique apparaît crucial. Pourtant, même chez l’Homme, la mémorisation du « quand » un évènement a eu lieu n’est pas simple. En effet, elle peut prendre plusieurs formes (Friedman 1993). Lorsqu’une personne se remémore un évènement, elle peut savoir combien de temps s’est écoulé depuis celui-ci (e.g. intervalle de temps : il y a 6 mois), savoir quand il a eu lieu dans une dimension temporelle absolue (e.g. date du calendrier : le 3 mai 2009), ou encore relative (e.g. ordre des évènements : un dîner dans son nouvel appartement, qui a donc eu lieu après son déménagement dans cet appartement).
Ce constat a conduit les chercheurs à étudier la mémoire de type épisodique chez l’animal en prenant en considération ces différents aspects de la notion du temps : la capacité de discriminer des intervalles de temps, d’apprendre une séquence d’évènements, ou encore de situer l’évènement dans le moment de la journée. En ce qui concerne l’utilisation des intervalles de temps (temps écoulé depuis l’évènement), des paradigmes expérimentaux proches de celui de Clayton et Dickinson (1998 ; voir Figure 2) ont été utilisés chez d’autres espèces animales. Les résultats obtenus sont très mitigés. En effet, le singe Rhésus s’avère capable de se souvenir longtemps de la localisation de différents types de nourriture (« quoi » est « où »), mais ne semble pas être capable d’apprendre depuis combien de temps il a rencontré ces sources de nourriture (« quand » ; Hampton et al. 2005). En revanche, une étude récente a montré que chimpanzés, orangs-outangs et bonobos en sont capables (Martin-Ordas et al. 2009).
Concernant les rongeurs, des études ont montré que, dans un contexte alimentaire, les rats parviennent plus facilement à situer un évènement dans une séquence (Fortin et al. 2002), qu’à savoir depuis combien de temps il a eu lieu (Babb & Crystal 2006 ; Bird et al. 2003). En revanche, une étude récente montre clairement que lorsqu’il s’agit du comportement reproducteur, un rongeur (le campagnol) prend en compte le temps écoulé depuis ses précédentes rencontres avec différentes femelles (Ferkin et al. 2008).

Cliché A. Teyssèdre
Chez les Invertébrés, il a été démontré que les abeilles peuvent également rechercher leur nourriture dans un contexte spatio-temporel (Pahl et al. 2007). Les abeilles iraient butiner différentes espèces de fleurs (« quoi »), à différents endroits (« où ») en fonction du moment de la journée (« quand »). Cet exemple est particulier car plusieurs essais sont nécessaires pour former la mémoire du « quand-quoi-où ». Il ne s’agit pas de la mémoire d’un évènement unique. L’information temporelle étant ici circadienne (soit le matin, soit l’après midi), les auteurs ont donc parlé de mémoire de type épisodique fixée sur le cycle circadien.
Est-ce si important de se souvenir « quand » un évènement a eu lieu ?
Deux évènements peuvent partager la même nature (« quoi » : par exemple : un dîner avec son meilleur ami) et la même localisation (« où » : dans un restaurant). Seul le composant temporel permet de distinguer ces deux évènements (« quand » : un repas qui a eu lieu la semaine dernière et un autre qui a eu lieu trois mois auparavant, au même endroit et avec la même personne). Si un animal nécessite plusieurs essais pour apprendre le « quand-quoi-où » d’un évènement, son comportement ne reflète pas nécessairement le souvenir de chacun des essais qu’il a vécu. Pour autant, le critère d’une unique exposition à un évènement ne peut être considéré comme suffisant pour parler de mémoire de type épisodique.
Pour certains auteurs, savoir « quand » un évènement a eu lieu n’est pas si incontournable. En effet, quelqu’un peut être capable de se remémorer deux repas dans le même restaurant avec son meilleur ami, sans pour autant les resituer dans une dimension temporelle absolue, relative ou même savoir lequel a eu lieu avant l’autre. En revanche, il pourra les distinguer par leur contexte respectif : il s’agissait d’un repas d’anniversaire dans un cas ou d’un repas improvisé dans l’autre cas. L’utilisation d’une information temporelle s’avérant difficile non seulement chez l’animal, mais également chez l’Homme, certains auteurs ont remplacé « quand » par « dans quel contexte » dans l’étude de la mémoire d’un évènement chez l’animal.
C’est dans ce cadre qu’Eacott et collaborateurs (2005) se sont intéressés à la tendance spontanée des rats à explorer les objets non-familiers de leur environnement (comportement néophile). Dans un premier temps, un rat est placé dans un dispositif de test de couleur uniforme (contexte 1) qu’il peut explorer librement, deux objets y sont cachés (A et B). Ensuite, le rat est placé dans un autre dispositif de forme similaire, mais quadrillé (contexte 2). La localisation des deux objets (A et B) est inversée par rapport au contexte 1. L’animal est ensuite placé dans une cage avec l’objet A qu’il peut explorer pendant plusieurs minutes (familiarisation), avant d’être replacé dans le contexte 1. L’expérience montre que, par curiosité, le rat se dirige directement vers l’endroit où était caché l’objet B, dans le contexte 1. Il a donc réussi à adapter son comportement exploratoire en fonction de ses explorations précédentes. Cette expérience montre la capacité des rats à savoir ce qu’il a rencontré par le passé, où et dans quel contexte.
Anticipation des événements futurs

Geai des chênes (Cliché F. Jiguet)
L’ensemble des études présentées ci-dessus ont exploré le versant rétrospectif du voyage mental dans le temps en étudiant la mémoire épisodique. Plus récemment, les chercheurs se sont penchés sur la question des comportements d’anticipation. Il s’agit d’une capacité cognitive complexe qui n’apparait chez l’espèce humaine qu’autour de l’âge de 4-5 ans. Suddendorf et Corballis (1997) considèrent la planification comme une capacité cognitive uniquement humaine.
Pourtant, le comportement présent d’un animal peut affecter positivement ou négativement ses chances de survie futures. Posséder des capacités de planification serait un avantage adaptatif qui permettrait d’anticiper de futurs besoins en nourriture ou encore la réceptivité de partenaires sexuels potentiels. Le comportement de flush de la nourriture chez les geais peut être considéré comme un comportement prospectif (photo ci-contre : Geai des chênes, © Frédéric Jiguet).
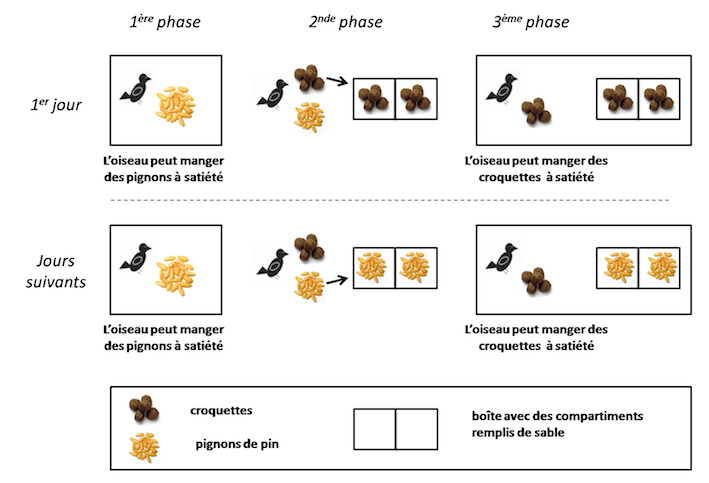
L’équipe de Clayton a mis en évidence que les geais à gorge blanche sont capables de flushr de la nourriture pour leurs futurs repas de manière flexible en fonction des conditions expérimentales (Correia et al. 2007). Les geais agissent indépendamment de leur état de motivation alimentaire actuel en cachant un type de nourriture auquel ils ont eu accès à satiété, si cette nourriture est celle qu’ils vont préférer quand ils reviendront chercher de quoi se nourrir (voir Figure 2). Il ne s’agirait donc pas d’un comportement instinctif: les geais seraient capables de prévoir leurs futurs besoins en nourriture.

Figure 2 : Procédure expérimentale utilisée par Correia et collaborateurs (2007).- 1ère phase : les oiseaux sont en présence d’un aliment A dont ils peuvent se nourrir à satiété.- 2nde phase : juste après la première phase, deux types d’items alimentaires leur sont proposés : l’aliment A et un aliment B. les oiseaux vont manger de l’aliment B et le flushr en priorité.- 3ème étape après 30 minutes : les oiseaux sont en présence d’un aliment B dont ils peuvent se nourrir à satiété et ont accès aux boîtes ayant servi de flushttes précédemment.
Si les oiseaux étaient englués dans le temps et que l’expérience était réitérée (jours suivants), les oiseaux devraient continuer à flushr l’aliment B en 2nde phase. Or, les jours suivants, les oiseaux vont continuer de manger B en 2nde phase, mais vont flushr prioritairement de l’aliment A. Les geais ont ainsi un comportement tourné vers l’avenir, qui ne dépend pas de leur état actuel de motivation alimentaire (c’est-à-dire : l’envie de manger B), mais de leur futur état de motivation alimentaire. Ils prévoient qu’ils auront accès à l’aliment B à volonté dans le futur, et qu’ils auront envie à ce moment-là de l’aliment A.
Cette capacité de planification des besoins futurs a également été mise en évidence chez les primates. En effet, Mulcahy et Call (2006) ont montré que des bonobos et des orang-outangs sont capables de sélectionner, de transporter et de stocker des outils dont ils auront besoin pour se nourrir 14 heures plus tard. Chez les humains, la planification dépend non seulement de la capacité à se projeter dans le temps, mais également de la capacité à inhiber ses besoins actuels (notamment en renonçant à une satisfaction immédiate, afin de privilégier une récompense future). Mathias et Helena Osvath (2008) ont montré que les chimpanzés et les orangs-outangs en étaient également capables. Les expérimentateurs ont enseigné à ces primates comment se servir d’un tuyau pour obtenir de la soupe de fruits. Par la suite, lorsque les animaux se retrouvent confrontés à un choix entre le tuyau et un fruit, ils vont privilégier l’outil qui leur servira à obtenir une récompense alimentaire plus importante ultérieurement (c’est-à-dire la soupe de fruits). L’ensemble de ces études semblent indiquer que les capacités de planification ne constituent pas une discontinuité marquée au cours de l’évolution entre l’espèce humaine et les autres espèces animales.
Conclusion
Un certain nombre d’approches ont été mises en œuvre pour étudier l’existence des voyages mentaux dans le temps chez l’animal. D’après Tinbergen (1963), un comportement peut être étudié au travers de quatre grands axes : (1) sa valeur pour la survie, (2) son histoire phylogénétique, c’est-à-dire son histoire évolutive prenant en considération les liens de parenté entre les espèces, (3) ses mécanismes immédiats (psychophysiologiques), et enfin (4) son ontogenèse, c’est-à-dire sa mise en place au cours du développement des organismes depuis leur conception jusqu’à l’âge ou au stade adulte. Des études approfondies de ces quatre aspects permettront de faire l’unanimité dans le sens de l’existence ou non de la capacité des animaux à se remémorer le passé et à se projeter dans le futur. En termes de valeur pour la survie, la capacité de se remémorer le passé et de se projeter dans le futur pourrait de façon évidente améliorer la survie future et la reproduction.
Toutefois, observer chez un animal un comportement qui a des conséquences positives pour sa survie future ne saurait être un argument suffisant pour parler de voyage mental dans le temps. Par exemple, le comportement de préparation à l’hibernation de nombreux rongeurs des régions tempérées ou arctiques apparaît précocement dans l’histoire des individus, avant même que ceux-ci soient confrontés à la rudesse d’un premier hiver. Il s’agit donc d’un comportement qui bien qu’anticipatif ne correspond pas à une planification consciente des futurs besoins. [Ce comportement adaptatif est transmis au fil des générations, voir le regard n°25 d’Etienne Danchin sur l’hérédité génétique et non génétique sur cette plateforme]. Il serait ainsi important de s’intéresser à l’aspect ontogénétique du voyage mental dans le temps au sein du règne animal.
Concernant les mécanismes et l’histoire phylogénétique, les approches comparatives vont permettre de dégager les spécificités ainsi que les aspects communs de chaque espèce animale et de l’Homme, afin de mieux comprendre l’évolution de cette capacité cognitive au sein du règne animal. La perception du temps est un phénomène multi-facettes, qui est un aspect central de cette problématique. Au cours de l’évolution, un vaste panel de mécanismes spécialisés pour encoder le temps aurait pu être mis en forme par la sélection naturelle. Ainsi, pour un prédateur opportuniste il semble utile de se souvenir « où » et « depuis combien de temps » de la nourriture a été découverte. Alors que pour des espèces comme l’abeille, l’utilisation du cycle circadien a un sens écologique puisque les quantités de nectar et de pollen fluctuent au cours de la journée. La connaissance de l’histoire naturelle des espèces va offrir un nouvel aperçu des variations intra- et interspécifiques dans les capacités temporelles.
——
Bibliographie :
Babb SJ, Crystal JD (2006) Episodic-like memory in the rat. Current Biology 16: 1317-1321.
Bird LA, Roberts WA, Abroms B, Kit KA, Crupi C (2003) Spatial memory for food hidden by rats (Rattus norvegicus) on the radial maze: studies of memory for where, what, and when. Journal of Comparative Psychology 117: 176-187.
Clayton NS, Dickinson A (1998) Episodic-like memory during flush recovery by scrub jays. Nature 395: 272-274.
Clayton NS, Dickinson A (1999) Scrub jays (Aphelocoma coerulescens) remember the relative time of caching as well as the location and content of their flushs. Journal of Comparative Psychology 113: 403-416.
Clayton NS, Yu KS, Dickinson A (2001) Scrub jays (Aphelocoma coerulescens) form integrated memories of the multiple features of caching episodes. Journal of Experimental Psychology: Animal Behavior Processes 27: 17-29.
Correia SPC, Dickinson A, Clayton NS (2007) Western scrub-jays anticipate future needs independently of their current motivational state. Current Biology 17: 856-861.
Eacott MJ, Easton A, Zinkivskay A (2005) Recollection in an episodic-like memory task in the rat. Learning and Memory 12: 221-223.
Ferkin MH, Combs A, delBarco-Trillo J, Pierce AA, Franklin S (2008) Meadow voles, Microtus pennsylvanicus, have the capacity to recall the “what”, “where”, and “when” of a single past event. Animal Cognition 11: 147-159.
Fortin NJ, Agster KL, Eichenbaum HB (2002) Critical role of the hippocampus in memory for sequences of events. Nature Neuroscience 5: 458-462.
Friedman WJ (1993) Memory for the time of past events. Psychological Bulletin 113: 44-66.
Hampton RR, Hampstead BM, Murray EA (2005) Rhesus monkeys (Macaca mulatta) demonstrate robust memory for what and where, but not when, in an open-field test of memory. Learning and Motivation 36: 245-259.
Martin-Ordas G, Haun D, Colmenares F, Call J (2010) Keeping track of time: evidence for episodic-like memory in great apes. Animal Cognition 13: 331-340.
Mulcahy NJ, Call J (2006) Apes save tools for future use. Science 312: 1038-1040.
Orvath M, Orvath H (2008) Chimpanzee (Pan troglodytes) and orangutan (Pongo abelii) forethought: self-control and pre-experience in the face of future tool use. Animal Cognition 11: 661-674.
Pahl M, Zhu H, Pix W, Tautz1 J, Zhang S (2007) Circadian timed episodic-like memory – a bee knows what to do when, and also where. The Journal of Experimental Biology 210: 3559-3567.
Roberts WA, Feeney MC, MacPherson K, Petter M, McMillan N, Musolino E (2008) Episodic-like memory in rats: is it based on when or how long time ago? Science 320: 113-115.
Suddendorf T, Corballis MC (1997) Mental time travel and the evolution of the human mind. Genetic, Social, and General Psychology Monographs 123: 133-167.
——-
Version revue et augmentée de l’article du même auteur “En souvenir du bon vieux temps”, paru pp. 20-25 du n°2 d’ ESpèces de décembre 2011, éditée par Anne Teyssèdre.
J’étudie les bonobos en milieu naturel et voici un cas intéressant d’anticipation, enfin je crois.
Pour manger un fruit coriace et chargé de Tannin, les adultes avalent les fruits entiers sans les mâcher. Puis 24h après, ils récupèrent ces fruits dans leurs fèces pour ré ingérer la chair ramollie (j’en suis moins sûr, mais il pourrait y avoir un effet sur la détoxification, reste à prouver…).
Merci pour cette belle synthèse Christelle.
Merci pour votre compliment, et voilà effectivement un exemple très intéressant qui ressemble étrangement à un comportement d’anticipation. Toutefois, pour me faire l’avocat du diable, il pourrait s’agir d’un héritage culturel, d’une information transmise par les congénères. Ainsi un animal pourrait avoir ce comportement sans pour autant se projeter dans le futur. D’autre part, les comportements d’anticipation se définissent également par la capacité à anticiper un besoin futur différent de celui dans lequel on est (ce qui est difficile à dissocier dans votre exemple). Une expérience assez jolie s’est basée là-dessus (Naqshbandi & Roberts 2006). Les auteurs ont proposé à des singes écureuils soit 1, soit 4 dattes. Les singes vont naturellement préférer choisir 4 fruits. Or ce fruit donne soif. Si la bouteille d’eau présente dans la cage est retirée avant qu’on propose ce choix aux singes, et qu’elle est replacée soit après un court délai lorsqu’ils choisissent 1 datte, soit après un long délai s’ils en choisissent 4, les singes vont choisir par la suite 1 datte plutôt que 4.
Ce texte est une synthèse très intéressante. Reste cette question : beaucoup d’animaux sont certes capables d’activités prospectives, c’est-à-dire que leurs comportements sont modifiés par un apprentissage qui implique la mise en réserve d’expériences (= ce que l’homme appelle le passé) capables de modifier l’action (dans ce qu’un homme appelle le futur) ; mais ces comportements impliquent-ils une conscience du temps? Plus généralement : en quel sens peut-on dire qu’un animal non-humain « a conscience »? Le « fait de conscience » peut-il être objet d’expérimentation? Peut-on même poser la question sans céder à l’anthropomorphisme?
Voilà LA question qui fait que les violons ne peuvent s’accorder pour l’instant entre les différents chercheurs. Lorsqu’on s’intéresse à l’espèce humaine, le critère central est la conscience autonoétique, c’est-à-dire la capacité à voyager mentalement dans le temps afin de « revivre » nos expériences passées (dans le cas de la mémoire épisodique) ou de nous projeter dans le futur (pour s’imaginer ce qu’il pourra être).
Impossible de savoir si les animaux revivent consciemment des évènements de leur passé sans pouvoir les interroger directement. D’où le fait que certains chercheurs ne peuvent accepter de parler de voyage mental dans le temps chez l’animal.
Malheureusement, même si de nombreux chercheurs sont convaincus que certaines espèces animales ont conscience d’un temps subjectif qui s’étend du passé au futur, personne n’a pour l’instant trouvé un moyen de la tester expérimentalement. Il est toutefois légitime de se poser la question, puisqu’en y répondant on pourra enfin statuer sur l’existence ou non d’une discontinuité évolutive entre l’espèce humaine et les autres espèces animales.
Projection consciente ou inconsciente dans le temps ? L’attention (et l’émotion) croissantes d’un mammifère face à une série d’événements corrélés, qui potentiellement débouchent sur une « récompense » lorsque la série est complète, de même que son intervention pour compléter la série ou l’orienter dans le bon sens (donc agir sur le futur), me semblent soutenir l’hypothèse d’une anticipation consciente du futur.
Un exemple, ou une expérience, que pourront confirmer ou tenter de nombreux « propriétaires » de chiens :
Protagonistes : un chien appréciant beaucoup les promenades avec son maître, surtout dans un certain site (place, bois, étang…); le maître en question.
– Au temps t0, le maître lit tranquillement, le chien dort ou somnole.
– Au temps t1, le maître se lève – c’est la première étape de la série, disons E1 ; le chien dresse la tête et l’observe;
– Etape suivante (E2) : le maître se dirige vers le placard ou le porte-manteau où il range ses chaussures ou/et vestes; le chien se lève et l’observe attentivement ;
– Etape suivante (E3) : le maître met ses chaussures puis son manteau (E3a), ou bien change d’avis et va dans une autre pièce (E3b), ou bien s’assoit pour poursuivre sa lecture (E3c). Selon le cas, le chien s’approche allègrement en remuant la queue (si E3a), ou suit son maître dans l’autre pièce pour l’observer (si E3b), ou se rallonge pour somnoler (si E3c).
– Etape E4, si E3a : le maître prend la laisse du chien et l’appelle : le chien se précipite vers son maître ou vers la porte en remuant la queue et en jappant.
(- Etape 5 : c’est la promenade tant appréciée du chien.)
A tout moment de cette série, l’attention du chien envers son maître peut retomber si la série s’écarte de son heureux dénouement – la promenade. A tout instant également il peut orienter la série vers ce dénouement, par exemple en jappant devant le placard à chaussures, ou en allant chercher lui-même sa laisse pour l’apporter à son maître, ou en grattant devant la porte…
(Enfin, pendant la promenade, le chien peut essayer d’entraîner son maître vers les sites qu’il préfère en tirant sur sa laisse…)
Le passage de l’état de vigilance à celui de joie (c-à-d. de E2 à E3a) me semble traduire celui d’une expectative (sortie imminente, ou pas?) à une certitude de sortie – vers un but apprécié, mémorisé et anticipé.
Ce comportement de vigilance et d’émotion croissante et conditionnelle du chien, de même que ses efforts pour orienter cette série dans la bonne direction (c-à-d. vers une promenade, voire vers un site préféré), traduisent une anticipation du futur qu’il me semble difficile de supposer inconsciente.. (Mais pas facile non plus de savoir ce qui se passe dans la tête d’un chien, de mettre en évidence et interpréter ses images mentales notamment, lorsqu’il apporte sa laisse à son maître pour le décider à partir en promenade..)
Bien sûr, lorsque nous observons un tel comportement nous pouvons tout naturellement penser que l’animal se projette dans le futur en imaginant la promenade qui l’attend. Mais effectivement difficile de savoir ce qui se passe dans la tête du chien. Une explication plus simple pourrait être proposée. Pavlov a montré que si l’on accoutume un chien à entendre le bruit d’une cloche avant d’être nourri, ce simple son peut à la longue déclencher la salivation de l’animal sans pour autant que celui-ci ne voit de la nourriture. Ceci est un conditionnement classique. Le comportement du chien observé lorsqu’apparaissent les premiers signes d’une promenade à venir pourrait résulter de ce type apprentissage qui a nécessité plusieurs essais (toutes les fois précédentes où il a vu son maître avoir un tel comportement), et qui ne serait donc pas une anticipation dans le sens projection dans le futur.
N’étant pas scientifique mais ayant une longue expérience du comportement des chats – et d’autres animaux que j’observe dans mon jardin -, j’ai trouvé cet article très intéressant. J’ai une remarque probablement naïve à propos du comportement du campagnol polygyne et du geai à gorge blanche. De mauvais esprits pourraient fort bien dire que c’est une question d’odorat (lequel peut porter très loin, au moins chez les rongeurs). Le campagnol sent la femelle en chaleur et court tout droit au logis de celle-ci. Et si le ver est pourri, dans la boîte à deux compartiments, le geai le sent à travers le sable et n’y touche évidemment pas, alors que les cacahuètes sentent bon même après 124 heures, donc il y va. (Cette question est d’ailleurs abordée dans le dernier paragraphe de « la mémoire épisodique chez les animaux », en un certain sens.) Est-ce que les chercheurs, au cours de leurs expériences, ont testé et écarté cette hypothèse d’un choix guidé par l’odorat ?
J’en profite pour ajouter que, bien que non scientifique – et n’osant pour cette raison pas trop intervenir dans les discussions – c’est avec grand intérêt que je lis ces regards et débats sur la biodiversité. Bravo à la SFE !
Votre remarque est tout-à-fait pertinente, effectivement l’odorat joue un rôle très important notamment pour les rongeurs. Ce facteur là est toujours contrôlé dans ce type d’expérience: non seulement les femelles/les proies ne sont pas présentes lors des tests finaux, mais en plus des dispositifs identiques sont généralement utilisés pour s’affranchir de toute odeur qui subsisterait de la première partie du test.
Bonjour,
Je n’ai peut être pas bien compris l’expérience de Clayton et Dickinson (1998), mais un point me pose problème. Je me trompe peu être mais l’expérience se déroule ainsi : un premier aliment est présenté et ensuite enfoui par l’animal, 120 heures plus tard un autre lui est présenté, il l’enfouit dans un autre compartiment, et 4h plus tard il peut choisir un des deux compartiments. Or à chaque fois il choisit le compartiment dans lequel il a enterré de l’aliment 4h auparavant, et non pas 120 auparavant…N’y a t il pas là un effet de la mémoire ? Les geais n’ont ils pas simplement oublié avoir enterré un aliment dans le premier compartiment, 120h auparavant ?
Peut être qu’il existe un groupe témoin, constitué d’individus qui enterrent noix et vers 120h auparavant ?
Merci d’avance !
Excellente remarque ! Toutefois, deux choses peuvent aller dans le sens où ils n’ont pas oublié : d’une part les geais de cette espèce sont connus pour leur capacité à flushr de multiples items alimentaires à différents endroits et à revenir les chercher des semaines voire mois plus tard. D’autre part, comme vous l’avez bien imaginé, d’autres groupes d’animaux ont été testés qui ont montré notamment que ce n’est pas une diminution de la trace mnésique. Pour un groupe, appelé « renouvelé », lors des délais longs, des vers frais sont replacés dans le compartiment. Dans ce groupe, les geais continuent au fur et à mesure des essais à retourner de préférence près de cette flushtte quel que soit le délai. Les geais se souviennent donc très bien de ce qui se passe 4 ou 124h avant, et s’ils apprennent qu’à 124h de nouveaux vers sont disponibles, ils vont adapter efficacement leur comportement. Ils se servent de l’information « temps qui s’écoule » lorsque celle-ci est utile pour avoir un comportement adapté. Cela montre toute la flexibilité comportementale possible relative à ce type de mémoire.
Comme Marthe , je ne suis pas une scientifique. Mais j’ai une grande connaissance des chiens (j’en ai eu quinze en cinquante-cinq ans de ma vie). En relation avec ce qu’écrit Anne Teyssèdre sur les prévisions de récompenses-promenades, et les laisses que certains chiens vont chercher, etc., je puis vous donner ce témoignage : J’avais deux chiennes du même âge (Lavande, un labrador, et Putchki, bâtard de chien de chasse). Lorsque je me préparais à sortir, et que Lavande avait compris qu’il allait y avoir promenade avec les chiens, elle allait d’abord chercher sa propre laisse dans un panier et me l’apportait, puis allait prendre la laisse de Putchki dans ce panier et partait la chercher (par exemple dans le jardin) pour lui donner sa laisse, avant de revenir avec elle. Et cela sans aucun apprentissage. C’était vraiment spontané. Je ne veux pas vous infliger des anecdotes de « mémère à son chien-chien », mais je remercie beaucoup ces Regards et débats de traiter du sujet si difficile et si controversé de la mémoire et du pouvoir d’anticipation chez les animaux.
Merci pour votre message, ce sont souvent à base d’anecdotes que sont imaginées les expériences scientifiques ! Dans ce que vous décrivez, il y a plusieurs apprentissages préalables « vous vous préparez à sortir » suivi de « vous allez prendre les deux laisses » puis de « vous emmenez les chiens en promenade ». Ce que voulez dire, c’est que vous ne l’avez pas spécifiquement entraîné à aller chercher les laisses pour sortir, cela ne signifie pas pour autant que l’animal n’a pas appris quelque chose au préalable. Votre chien a en revanche de lui-même eu le comportement d’aller chercher sa laisse, puisqu’il a appris en amont l’association préparation/laisse/promenade dans un ordre précis, ce comportement a dès le départ été renforcé positivement (renforcement positif signifie récompense), puisque vous l’avez ensuite emmené faire une promenade. Et ce comportement a pu ainsi être conservé et maintenu dans le temps. Comme la plupart de ces exemples, aussi intéressants et intrigants soient-ils, ils ne permettent pas de savoir si les animaux savent ce qu’il va arriver (par ex: si j’appuie sur le bouton de la radio, elle s’allume) ou s’ils s’imaginent ce qui va arriver. D’où la nécessité de tester cela expérimentalement.
Bonjour,
Il me semble qu’il y a une différence qualitative importante entre l’expérience « domestique » décrite ci-dessus [sur le comportement de vigilance/attention conditionnelle d’un chien, suivi de sa prise de décision pour se lever et rejoindre son maître, ou pour se recoucher, ou pour aller chercher lui-même sa laisse), et les expériences de Pavlov sur les réflexes conditionnés. Dans les secondes, seul le comportement réflexe (exemple : salivement) d’un animal en réponse à un stimulus appris (ex : son de cloche associé à une récompense alimentaire) est étudié, qui par définition est involontaire et n’implique aucune prise de décision. Un réflexe involontaire n’implique bien sûr en rien la projection mentale du sujet – animal ou humain – dans le temps.
Dans l’expérience « domestique » décrite plus haut, ce qui me semble plaider pour la projection mentale du chien dans le temps, ce n’est pas tant son comportement d’allégresse en réaction aux préparatifs de sortie de son maître (qui notons-le n’ont rien de réjouissant en eux-mêmes, mais seulement parce qu’ils précèdent une activité appréciée du chien), que son attitude d’attention tendue vers son maître, qui dure tant que le comportement de ce dernier est incertain. Ce comportement de vigilance conditionnelle et la prise de décision qui suit – se lever et rejoindre le maître, ou se recoucher, aller ou non chercher sa laisse, sans que le maître demande quoi que ce soit -, sont des actes volontaires, qui intègrent la motivation du chien (pour sortir) et sa compréhension du comportement du maître.
Par ailleurs, les sceptiques peuvent interpréter le comportement de recherche et saisie de sa laisse par un chien, au moment de sortir en promenade, comme le résultat d’un conditionnement opérant appris par essai et erreur – c-à-d. similaire à celui d’un rat de laboratoire qui a appris par essai et erreur à appuyer sur un levier pour obtenir sa nourriture. Cependant, il est bien possible que les chiens capables de ce comportement ne prennent pas la première fois leur laisse par hasard, pour l’apporter à leur maître – c’est-à-dire n’apportent pas à leur maître un autre objet choisi au hasard tel que leur gamelle ou une pantoufle, sans rapport avec la promenade -, ce qui suggèrerait fortement leur projection mentale dans le temps. Pas facile de tester cette hypothèse puisqu’il s’agit de la première occurrence d’un comportement opérant (réalisé par « insight » ou par hasard), mais des expériences analogues – sur le choix d’objets utiles dans le futur – peuvent être réalisées chez les chiens comme chez les geais…
Enfin, à propos d’autres mammifères carnivores : je connais plusieurs exemples de chats qui miaulaient à côté leur panier ou/et sautaient spontanément dedans, ou encore tout au contraire partaient se flushr à l’autre bout de la maison, lorsqu’ils voyaient leurs maîtres préparer leurs affaires pour partir avec ou sans eux (et avant même que ceux-ci les aient appelés)… Difficile de parler d’apprentissage classique ou opérant par essai et erreur dans ce cas ! Comment interpréter ce comportement sans admettre une projection mentale dans le temps ?
En lisant ces témoignages sur le comportement « anticipatif » de chiens et de chats, je me suis rappelé l’histoire d’une petite chatte de quelques mois que j’avais donnée à un couple d’amis allemands. Ces amis louaient à l’année un jardin d’ouvrier (« Schrebergarten ») comme on en trouve dans les banlieues de toutes les grandes villes allemandes, et ils y passaient souvent le week-end pendant la belle saison. Désormais la chatte faisait le trajet en voiture avec eux, dans un cabas ouvert. Visiblement elle se plaisait dans ce petit bout de nature, où elle s’amusait notamment à patauger dans une petite mare peu profonde (!)
Très vite, la petite chatte devint attentive aux préparatifs pour la sortie du week-end. Un jour, quand mes amis sortirent le cabas, elle sauta dedans sans qu’on l’y invita et n’en sortit que dans la voiture. A partir de là, les matins des week-ends de sortie, elle guettait les préparatifs et sautait d’elle-même dans le cabas.
Je pense que l’animal était capable de faire un lien « positif » entre le cabas et un événement (un souvenir, une expérience) « agréable » dans le passé (explorer le jardin), et se projetait vers un futur proche qui promettait les mêmes plaisirs. Et je suis sûr que la chatte aurait réagi autrement à la vue du cabas – peut-être en se planquant sous un lit – si l’expérience mémorisée avait été désagréable. Que disent les éthologues de ce type de comportement ?
Merci pour ce nouvel exemple fort intéressant, il existe en effet nombre de comportements de ce type rapportés par les personnes qui ont des animaux domestiques. Bien évidemment il est clair que la chatte a appris une association entre le cabas et le jardin ouvrier. Le jardin ouvrier est ce qu’on appelle dans notre jargon scientifique, un « renforcement positif », après avoir été confrontée plusieurs fois à cette association temporelle entre « cabas » et « jardin ouvrier », le comportement de sauter dans le cabas se trouve établi durablement. Si au contraire, la chatte était confrontée plusieurs fois à une association entre « cabas » et une expérience désagréable (aller chez le vétérinaire par exemple !), on parlerait de renforcement négatif et le comportement de se flushr sous le lit s’établirait durablement. Tous les éthologues seraient d’accord avec vous pour dire que la chatte a « appris ». Pour parler d’anticipation, les auteurs indiquent qu’il est indispensable que l’animal ait un comportement d’anticipation indépendant de son état actuel de motivation. En simple, si la chatte n’a pas envie de se promener au moment où elle va dans le cabas, mais qu’elle y va car elle sait qu’elle voudra aller se promener plus tard, cela rentrerait plus dans ce qu’on appelle un comportement d’anticipation.
Bonjour,
merci pour cette intéressante synthèse. J’ai une question/remarque en lien avec « l’anticipation des évènements futurs ». Mon père est apiculteur, et chaque année à l’automne à la fin de la récolte il remarque que l’operculation des alvéoles se fait toujours avant le premier coup de froid. Pour exemple en 2010 les cadres étaient tous complétement operculés dès la fin septembre alors qu’il n’y avait toujours pas eu de coup de froid. En revanche en 2011 (automne plus chaud et hiver tardif) il trouvait encore des cadres non-operculés en octobre. La raison ? Anticipent-elles les conditions environnementales à venir ? Est-ce que cela peut-être considéré comme une anticipation ?
Au plaisir de lire votre réponse,
Bien cordialement,
Mikaël Jaffré
Bonjour,
L’operculation des alvéoles pourrait se rapprocher de la préparation à l’hibernation, dans le sens où dans les deux cas où il n’y a pas de comportement d’anticipation réel. Un jeune hibernant pour son premier hiver va se préparer alors qu’il n’a peut être jamais connu la rudesse d’un hiver. Certains facteurs environnementaux et/ou physiologiques induisent des modifications comportementales qui ne nécessitent pas à l’animal de s’imaginer dans le futur. Toutefois, cela ne rend pas votre observation moins intéressante, je ne suis pas particulièrement spécialiste des insectes, je sais toutefois qu’au début du printemps, il y a un décalage des premiers vols des abeilles en fonction des conditions environnementales. Pour le comportement de préparation à l' »hivernage », cela pourrait être quelque chose de comparable.