
La Société Française d’Ecologie (SFE) vous propose cette semaine le regard d’Etienne Danchin, chercheur au CNRS en écologie évolutive, sur l’hérédité non génétique et l’évolution.
MERCI DE PARTICIPER à ces regards et débats sur la biodiversité en postant vos commentaires et questions après cet article. Les auteurs vous répondront et une synthèse des contributions sera ajoutée après chaque article.
L’hérédité non génétique,
un changement de paradigme en sciences de l’évolution
Etienne Danchin,
Directeur de Recherche au CNRS, Université Paul Sabatier,
Laboratoire Évolution & Diversité Biologique
Regard R25, édité par Anne Teyssèdre
Mots clés : évolution, hérédité, espèces, populations, hérédité non-génétique, génétique, épigénétique, culture animale.
———
Une propriété majeure du vivant est la reproduction, à tel point qu’on distingue les organismes vivants des entités inanimées par cette propriété. Et une des caractéristiques fondamentales de la reproduction est que les organismes engendrent des descendants qui leur ressemblent. Cette ressemblance comporte deux composantes distinctes.

Cliché Eckard Berberich
Tout d’abord, les organismes descendants appartiennent à la même espèce que l’organisme d’origine. Cette propriété s’appelle l’hérédité. L’expression du langage courant « un chat ne fait pas un chien » la résume très bien, même si elle est souvent employée dans un contexte différent.
D’autre part, cette ressemblance s’exprime aussi de manière interne à l’espèce: les enfants ressemblent à leurs parents. En d’autres termes, les variations morphologiques, physiologiques et comportementales que l’on trouve au sein de la génération des parents se retrouvent au sein de la descendance et ce de manière associée de telle sorte que, par exemple, les parents de grande taille ont en moyenne des enfants de plus grande taille que celle des enfants de parents petits. Cette propriété fondamentale est appelée l’héritabilité.
C’est cette deuxième propriété qui a conduit Charles Darwin et Alfred R. Wallace à proposer le processus d’évolution par sélection naturelle. Ils ont en effet compris que la sélection naturelle ne pouvait conduire à l’évolution d’un trait que si les parents transmettaient leurs caractéristiques à leur descendance.
Prenons un exemple
Imaginons un éleveur qui vivrait de sa production de lait et aurait hérité de ses parents d’une étable dans laquelle il peut mettre vingt vaches. Imaginons aussi que son espace soit contraint et qu’il ne puisse pas agrandir son étable. Le seul moyen pour lui d’augmenter ses revenus serait d’avoir des vaches produisant plus de litres de lait par jour.

Cliché Anne Teyssèdre
Pour ce faire, il va, comme ses ancêtres depuis plus de 10 000 ans, choisir pour la reproduction les vaches qui font beaucoup de lait. Ce faisant, il exercera une forte pression de sélection en faveur des vaches produisant plus de lait que la moyenne, et la génération suivante ne contiendra que des descendantes de vaches faisant beaucoup de lait.
Le point important est que l’objectif d’augmenter la production de lait par vache au cours des générations ne se réalisera que si celles qui font beaucoup de lait ont des descendantes qui leur ressemblent sur ce caractère, autrement dit si le caractère « quantité de lait produite à l’âge adulte » est héritable (selon la définition plus haut). Sinon, quelle que soit la pression de sélection exercée par l’éleveur, la quantité de lait produite par jour n’augmentera pas de génération en génération. En d’autres termes, pour qu’une évolution sur un trait donné (ici la quantité de lait produite par jour par une vache) se produise suite à une sélection, il faut que ce trait soit héritable, c’est-à-dire que les caractéristiques individuelles des parents soient au moins en partie transmises aux descendants.
Hérédité et génétique
Bien entendu, en lisant ces lignes, je suis certain que nombre d’entre vous pensez que cette propriété d’héritabilité est due aux variations génétiques existant entre les vaches. C’est vrai en grande partie, mais c’est assez réducteur. Nous savons tous aujourd’hui que l’information génétique est universellement codée, au sein de chaque cellule, dans les séquences d’ADN (acide désoxyribonucléique), macromolécule semblable à une double chaîne comprenant des millions de maillons, chacun constitué d’une paire de bases parmi quatre différentes (désignées par les lettres A, T, G et C). C’est la succession de ces bases qui encode l’information génétique conduisant, entre autres, à la synthèse des protéines, molécules fondamentales de la constitution des cellules et de leur métabolisme.
© ATM2
Constituant des chromosomes, l’ADN se duplique à chaque division cellulaire, permettant ainsi la transmission fiable de l’information génétique de génération en génération. Bien que très fiable, cette duplication conduit régulièrement à des erreurs de copie, c’est-à-dire à des changements dans les séquences d’ADN, dits mutations. Celles-ci sont responsables de la variation génétique que l’on observe dans toutes les populations naturelles d’organismes vivants. Lorsque l’on parle de variation génétique, on sous-entend donc des variations dans la séquence de l’ADN entre organismes au sein d’une population. Nous pensons pour la plupart que l’hérédité et l’héritabilité sont uniquement dues à la transmission de gènes, et donc de leurs variations, de parent à descendant. Cette vision pourtant assez récente est devenue tellement classique qu’elle fait partie de la sagesse populaire d’aujourd’hui.
Cependant, cette vision de l’hérédité est remise en cause depuis plusieurs décennies ((Mameli, 2004; Bonduriansky & Day, 2009) et devient de moins en moins tenable aujourd’hui (Danchin et al., 2011). Cette remise en cause ne porte pas sur la réalité de la transmission génétique, qui est indiscutable, mais sur la réduction à cet unique mode de transmission. La fascination exercée par la transmission génétique nous a conduit à réduire l’hérédité à sa seule composante génétique.
Pour bien comprendre de quoi il retourne, il nous faut revenir à la nature profonde de l’hérédité. Qui dit reproduction implique un transfert d’information de la génération des parents vers les descendants. En quelque sorte, la reproduction implique que des instructions pour construire la descendance sont transmises des parents vers leurs descendants. L’hérédité et l’héritabilité sont dues au fait qu’il existe un (ou des) processus de duplication de l’information, processus se produisant dans le cadre de la reproduction. Une part conséquente de cette information est en effet encodée dans la séquence de l’ADN, c’est indiscutable. On doit d’ailleurs considérer les gènes non pas comme des morceaux d’ADN, mais plutôt comme des unités d’information et la sélection ne se fait pas sur l’ADN proprement dit mais bien sur l’information véhiculée par la séquence de l’ADN (Gilddon & Gouyon, 1989). Cette vision de la génétique met les concepts d’information biologique au centre de la biologie évolutive moderne.
L’hérédité non génétique
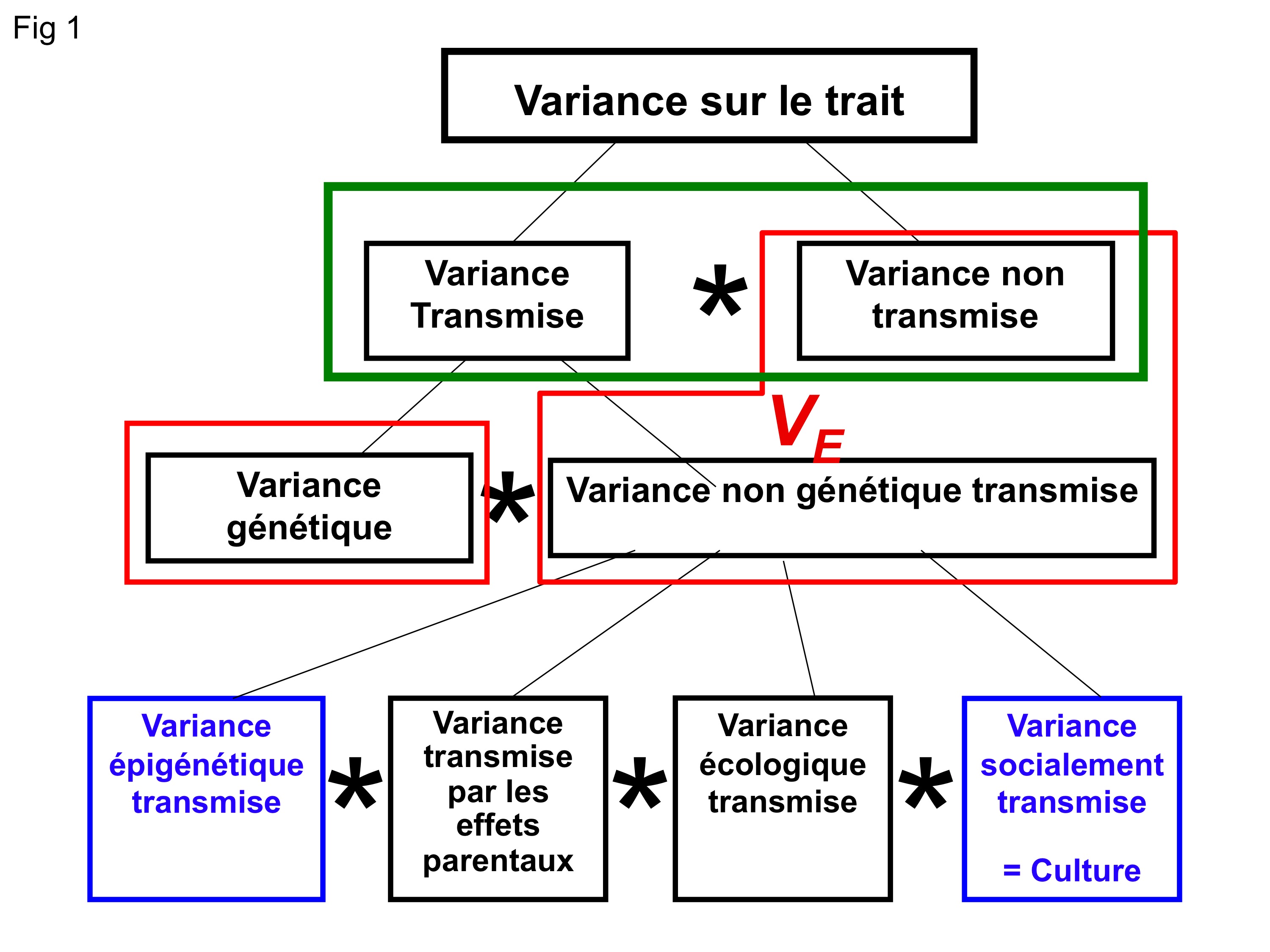
Fig. 1: Décomposition de la variance identifiant les différentes composantes de l’hérédité, qu’elles soient génétiques ou non. En rouge la décomposition généralement utilisée actuellement, qui nous conduit à penser que seul l’information génétique affecte l’évolution. En vert la décomposition que nous suggérons et qui correspond en fait à la vision initiale de Darwin. Celle-ci laisse la place à la prise en compte de l’hérédité non génétique. En bleu les deux types d’hérédité non génétique développés ici. Les étoiles indiquent que toutes ces composantes sont susceptibles d’interagir fortement.
Cependant, cette découverte majeure de la première moitié du XXième siècle qu’a été la génétique nous a tellement fascinés qu’elle nous a conduits à négliger toute autre forme de transfert d’information (et donc d’hérédité) entre les générations. Pourtant, il existe bien des formes de transfert d’information entre générations non basées sur la seule séquence de l’ADN. On parle alors d’hérédité non génétique, expression qui, aux vues de notre savoir collectif, semble associer deux concepts incompatibles.
Les découvertes qui s’accumulent depuis plusieurs décennies ont montré que ces transferts d’informations non génétiques entre générations sont bien réels et peuvent en fait prendre plusieurs formes très variées. Nous en avons récemment fait une revue et proposé un formalisme en terme d’hérédité dans Nature Reviews, Genetics (Danchin et al., 2011). En voici un bref aperçu en se focalisant sur deux formes d’hérédité non génétique parmi les quatre que nous avons identifiées (Figure 1).
Les deux autres formes d’hérédité non-génétique, que nous n’explorerons pas ici, sont celles résultant des effets parentaux par lesquels les parents peuvent mouler le phénotype de leur descendance dans les conditions environnementales qui prévalent, et l’hérédité écologique qui résulte du fait que les descendants héritent souvent de l’habitat des parents. Cette dernière forme d’hérédité crée des corrélations entre le génotype et l’environnement qui peuvent faire croire à une association adaptative entre gène et environnement. Elle peut aussi contribuer à ce qu’on appelle la « construction de niche », dans laquelle les activités des individus, en modifiant de manière durable les caractéristiques de l’environnement, changent les pressions de sélection sur la descendance.
L’hérédité épigénétique
L’épigénétique concerne l’étude de toute variation dans l’expression des gènes indépendamment de toute variation dans la séquence de l’ADN. C’est depuis cinq ans un domaine en pleine expansion et des découvertes très variées s’accumulent. Le point de départ est à trouver dans l’étude de la différenciation cellulaire au sein des organismes pluricellulaires. Cette différentiation n’est pas évidente à comprendre car toutes les cellules d’un organisme ont la même information génétique. Pourtant elles prennent des formes très différentes – pensez par exemple à des cellules d’os, ou de muscle, ou à des neurones. Ces variants de types cellulaires résultent de variations dans l’expression des gènes. Une image simple serait de comparer les gènes à des recettes: il est clair que deux personnes utilisant le même livre de recettes vont faire des repas très différents selon qu’elles utilisent telle ou telle recette. Il en est de même entre lignées cellulaires au sein d’un organisme pluricellulaire.
On s’est récemment rendu compte que des variations entre individus que l’on croyait due à des variations dans la séquence de l’ADN (les mutations responsables de la variation génétique) sont en fait dues uniquement à des variations dans l’expression de gènes qui sont identiques en termes de séquence d’ADN.

Linaire vulgaire, Cliché Noëlie Maurel
L’exemple classique est celui des fleurs péloriques de la linaire vulgaire, une fleur qui pousse sur les murs de nos villages. La fleur normale de linaire forme un tube constitué des pétales soudés à leur base, le tout ressemblant à une petite gueule de loup avec une symétrie bilatérale. Le grand botaniste et père de la systématique Carl von Linné fut surpris de trouver dans la descendance de ces plantes qu’il cultivait des fleurs à symétrie axiale avec cinq pétales bien distincts. Étant fixiste, il se trouvait confronté à une énigme insoluble car tout se passait comme s’il venait d’assister à la naissance d’une nouvelle espèce. Ce n’est que 250 ans plus tard qu’il a été démontré que ce cas apparemment typique de mutation n’était en fait pas dû à une mutation, la séquence étant identique entre les formes pélorique et normale, mais relevait d’un changement dans l’expression d’un seul gène (Cubas et al., 1999). Cette découverte fit grand bruit et fut publiée dans la revue Nature en 1999. Un autre exemple de ce type est développé dans la Figure 2.
Il apparaît aujourd’hui, de manière très surprenante, que de nombreuses variations épigénétiques sont transmises à la descendance. Les descendantes des plantes à fleurs péloriques découvertes par Linné il y a 250 ans ont elles aussi des fleurs péloriques et continuent de produire des descendantes à fleurs péloriques, démontrant ainsi l’existence d’une véritable hérédité non liée à des variations dans la séquence de l’ADN. Tout se passe comme si cette variation était due à une mutation, mais ce n’est pas le cas. Il existe donc bien une hérédité épigénétique.
Cette variation d’expression de certains gènes est due à divers mécanismes moléculaires dont le plus connu est celui de la « methylation » de la molécule de l’ADN, c’est-à-dire l’apposition répétée du radical méthyl (-CH3), un petit groupe de quatre atomes, sur une séquence de bases de l’ADN. Certaines conditions environnementales conduisent à la forte methylation de certains gènes spécifiques, réduisant, voire annulant l’expression de ces gènes, affectant ainsi la morphologie ou le comportement de l’individu concerné. De plus, lors de la duplication de l’ADN, il existe toute une batterie de mécanismes recopiant le pattern de methylation de l’ADN. C’est pourquoi les cellules filles sont généralement du même type que la cellule mère (cellules de foie, de muscle, etc.). De même, le cas des fleurs péloriques développé ci-dessus montre que les changements épigénétiques peuvent conduire à des situations où des caractères influencés par l’environnement sont transmis à la descendance sur de nombreuses générations! C’est tout à fait surprenant et remet en cause un des grands principes de la vision génocentrique que nous avons actuellement de l’évolution.
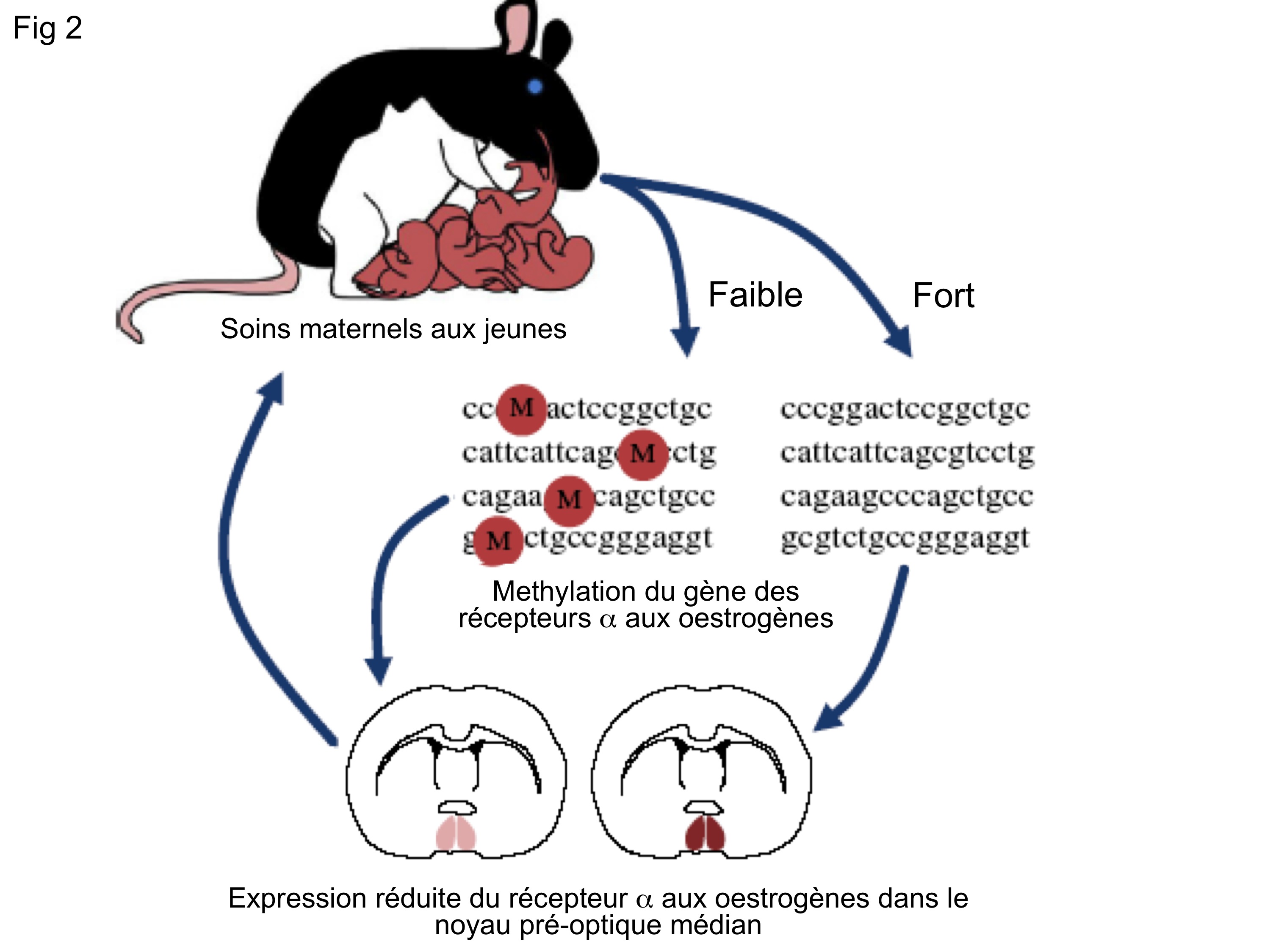
Fig. 2 : Hérédité épigénétique des soins maternels chez les rongeurs.
Les exemples d’une telle hérédité épigénétique s’accumulent actuellement tant chez les plantes que les animaux ou les microorganismes, où ils semblent jouer un grand rôle dans l’adaptation des organismes à leur milieu. L’intensité des soins maternels aux jeunes, chez les rongeurs, a ainsi une composante épigénétique comme le montre la figure 2 ci-contre (dans lequel les radicaux méthyl sont figurés par la lettre M).
Figure 2 : Hérédité épigénétique des soins maternels aux jeunes chez les rongeurs. Chez les rongeurs, les mères s’occupent intensément de leurs jeunes. Si l’on empêche expérimentalement des femelles de soigner leur progéniture, leurs filles à l’âge adulte délaissent leurs petits. Il a été démontré que cela implique la méthylation et donc la très faible expression du gène codant pour le récepteur alpha aux oestrogènes dans le cerveau des jeunes femelles issues de femelles les ayant peu soignées. A l’âge adulte, n’ayant que très peu de récepteurs aux oestrogènes dans leur cerveau, ces femelles de 2e génération sont insensibles à leurs propres œstrogènes circulant et de ce fait ne développent pas un comportement maternel normal (modifié de Champagne, 2008).
Une conclusion s’impose: si l’on se base sur les seuls patrons de transmission, il est facile de conclure à une transmission génétique, alors qu’il s’agit en fait d’hérédité des marques épigénétiques responsables de la variation entre individus, ou encore d’autres modes de transmission non génétique de l’information entre générations.
L’hérédité culturelle
Une autres formes d’hérédité non génétique est l’hérédité culturelle, dans laquelle l’information qui affecte l’aspect individuel (ou phénotype) est transmise socialement des anciens vers les plus jeunes. Jusqu’à récemment l’influence des sciences sociales humaines avait limité l’étude du phénomène culturel à la seule espèce humaine. Cependant, aujourd’hui force est de constater que la transmission sociale existe chez de très nombreux vertébrés et invertébrés où elle peut prendre des formes très variées comme l’imitation, le copiage, l’apprentissage social ou l’imprégnation sociale…
Par exemple, il a été démontré chez de nombreux animaux que les préférences sexuelles sont influencées par l’expérience sociale antérieure, affectant ainsi l’évolution génétique. Ainsi, chez certains poissons (Molly-voile, Poecilia latipinna) la préférence naturelle des femelles va aux mâles de grande taille. Cependant, on peut facilement inverser cette préférence en manipulant les informations sociales apportées aux femelles. Après avoir observé des petits mâles en train de copuler (donc apparemment attractifs pour les autres femelles) et des grands mâles seuls (et donc apparemment peu attractifs), ces femelles observatrices vont changer de préférence et choisir de petits mâles pour partenaires sexuels. On constate donc qu’un trait comportemental aussi important que la préférence sexuelle, trait que l’on croyait sous fort contrôle génétique, est en fait sous forte influence sociale, suggérant l’existence d’une véritable transmission sociale entre générations. De tels phénomènes ont été décrits chez des groupes aussi variés que des poissons, des oiseaux ou même des drosophiles.
Des expériences de ce type ouvrent tout un champ nouveau pour l’étude de l’hérédité. On peut en particulier imaginer que dans la nature des populations puissent diverger parce que des contingences locales (comme la présence de prédateurs préférant les gros individus) ont conduit les femelles d’une population à transmettre culturellement une préférence pour les petits mâles alors que dans d’autres populations en l’absence de tels prédateurs les femelles transmettent culturellement une préférence pour les grands mâles. Une telle différence va exercer des pressions de sélection sexuelle opposées dans ces populations les entraînant dans des chemins évolutifs qui à termes peuvent se révéler si différents que cela conduit à l’arrêt de tout transfert de gènes, ouvrant ainsi la voie vers la spéciation.
De nouveau, il apparaît que si l’on se base uniquement sur les patrons de transmission, il est facile de confondre la transmission sociale avec une transmission génétique. Un cas exemplaire est celui des poissons cichlides du lac Victoria, espèces où les parents s’occupent de leurs jeunes pendant le développement, offrant ainsi la possibilité d’imprégnation sociale. Certaines espèces jumelles sont si proches qu’il est difficile de les différencier. Pourtant, bien que ces poissons se reproduisent en colonies mixtes, très peu d’hybridations sont observées, ce qui pose la question du mécanisme de reconnaissance spécifique. Une première étude des préférences sexuelles avait conduit à conclure que cette préférence est « codée par un petit nombre de gènes sans dominance » (Haesler & Seehausen, 2005). Pourtant, deux ans plus tard, une expérimentation dans laquelle les alevins ont été échangés très tôt entre nids des deux espèces a montré que les jeunes apprenaient à reconnaître les membres de leur espèce lors du développement (Verzijden & ten Cate, 2007).
Une vision plus complète de l’hérédité
Il apparaît que l’hérédité non génétique peut prendre des formes variées. C’est l’interaction entre ces divers mécanismes de transmission d’information entre les générations qui façonne l’hérédité (Figure 3). Il n’est aujourd’hui plus possible de comprendre l’hérédité et donc l’évolution sans intégrer ces divers processus au sein d’une théorie synthétique moderne plus générale.
Historiquement, la synthèse moderne de la théorie de l’évolution désigne la synthèse qui s’est faite pendant la première moitié du XXième siècle entre la vision Darwinienne de l’évolution par sélection naturelle et la génétique. Aujourd’hui, de nombreux auteurs appellent à une nouvelle synthèse moderne intégrant toutes les formes d’hérédité, qu’elles soient génétiques ou non. Une vision plus complète de l’hérédité devrait nous permettre de résoudre de grosses énigmes évolutives. Par exemple, une importante énigme de la biologie moléculaire d’aujourd’hui réside dans le fait que la variation globale de la séquence de l’ADN analysée sur tout le génome ne permet d’expliquer que 5 à 10% de l’héritabilité de traits aussi variés que la taille corporelle des humains ou bien des maladies dites génétiques (Maher, 2008). C’est là une énigme fondamentale pour la médecine et le développement de thérapies adaptées.
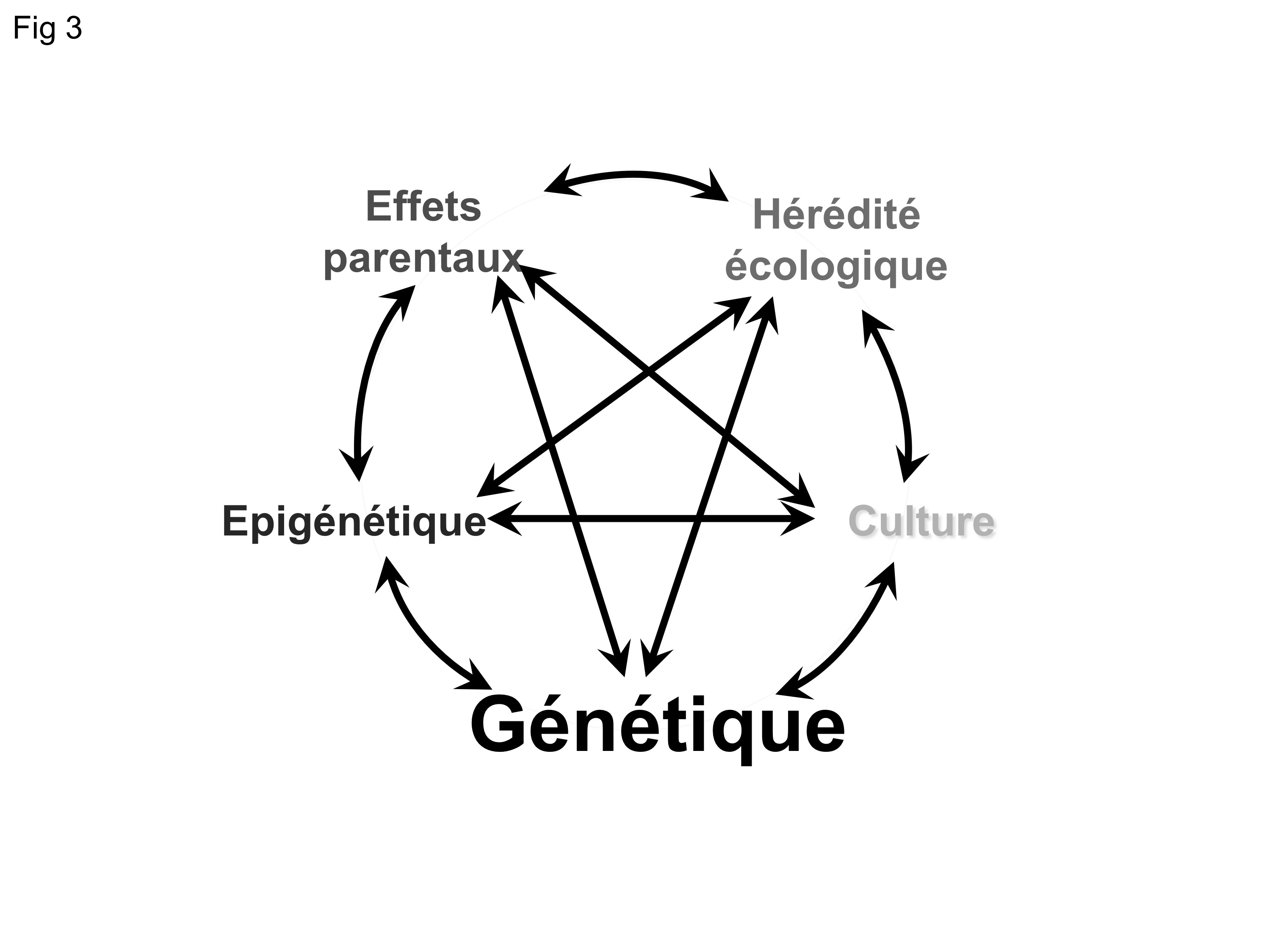
Fig. 3 : Tous les systèmes d’hérédité interagissent fortement entre eux pour produire l’hérédité. En négligeant l’hérédité non génétique, nous ignorons toute la richesse de ces interactions et des conflits que cela peut générer. En conséquence, nous manquons toute une gamme de dynamiques évolutives complexes qui pourtant participent à l’évolution.
Il est important de souligner que ces divers mécanismes d’hérédité ont des propriétés très différentes, en particulier en termes de transmission. Par exemple, alors que chez les organismes dont les cellules ont des noyaux (dits « eucaryotes) la transmission génétique ne se produit que « verticalement », de parent à progéniture, la transmission culturelle se fait aussi « horizontalement », c’est-à-dire entre individus de la même génération. Ainsi l’information héritable suit des chemins très différents selon la nature des mécanismes qui la sous-tendent. L’information génétique ne peut se transmettre que verticalement et donc mettra de nombreuses générations pour envahir toute une population; en comparaison, l’information culturelle peut, se répandre beaucoup plus rapidement et peut théoriquement envahir une population en une seule génération. Ces différentes propriétés ont d’importantes conséquences sur les dynamiques adaptatives et évolutives. Ainsi, des processus qui sont réputés impossibles avec une transmission verticale (comme c’est le cas de la transmission génétique) deviennent possibles si l’on inclut des processus culturels par exemple.
Ces propriétés particulières de la transmission non génétique ont conduit certains auteurs à faire remarquer que cela ouvrait la possibilité d’une forme d’hérédité de caractères acquis par les parents pendant le développement, ce qui a « un parfum délicieusement lamarckien » (Bird, 2007).
Conclusion
La conclusion, c’est que l’hérédité non génétique a pour effet de changer en profondeur les règles de fonctionnement de l’évolution et peut affecter amplement la transmission génétique. Des études récentes ont ainsi montré que des centaines de gènes humains ont récemment été sous forte sélection positive, souvent en réponse aux activités humaines, et que la culture humaine peut façonner le génome humain en profondeur (Laland et al., 2010).
Cette nouvelle vision nous conduit à redéfinir le processus d’évolution. Classiquement l’évolution se définit comme « le processus par lequel les fréquences de gènes changent au cours des générations ». Cette définition par trop réductrice devrait être remplacée par une définition plus large même si elle ne diffère que par un seul mot: « l’évolution c’est le processus par lequel les fréquences de variants changent au cours des générations » (Bentley et al., 2004). Par variant nous entendons bien entendu les gènes mais aussi tout type d’information transmis de génération en génération et participant donc à l’évolution des espèces.
Pour faire court, on peut dire que nous vivons aujourd’hui un véritable changement de paradigme dans les sciences de l’évolution, changement si profond que l’on peut se hasarder à prédire que la période actuelle sera plus tard perçue comme une véritable révolution conceptuelle.
Bibliographie
Bentley, R. A., Hahn, M. W. & Shennan, S. J. 2004. Random drift and culture change. Proceeding of the Royal Society of London B, 271, S353-S356.
Bird, A. 2007. Perceptions of epigenetics. Nature, 447, 396-398.
Bonduriansky, R. & Day, T. 2009. Nongenetic Inheritance and Its Evolutionary Implications. Annual Review of Ecology, Evolution, and Systematics, 40, 103-25.
Champagne, F. A. 2008. Epigenetic mechanisms and the transgenerational effects of maternal care. Frontiers in Neuroendocrinology, 29, 386-97.
Cubas, P., Vincent, C. & Coen, E. 1999. An epigenetic mutation responsible for natural variation in floral symmetry. Nature, 401, 157-161.
Danchin, É., Charmantier, A., Champagne, F. A., Mesoudi, A., Pujol, B. & Blanchet, S. 2011. Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nature Reviews Genetics, 12, 475-486.
Gilddon, C. J. & Gouyon, P. H. 1989. The units of selection. Trends in Ecology and Evolution, 4, 204-208.
Haesler, M. P. & Seehausen, O. 2005. Inheritance of female mating preference in a sympatric sibling species pair of Lake Victoria cichlids: implications for speciation. Proceedings of the Royal Society of London, 272, 237-245.
Laland, K. N., Odling-Smee, J. & Myles, S. 2010. How culture shaped the human genome: Bringing genetics and the human sciences together. Nature Reviews Genetics, 11, 137-148.
Maher, B. 2008. Personal genomes: The case of the missing heritability. Nature, 456, 18-21.
Mameli, M. 2004. Nongenetic selection an nongenetic inheritance. British Journal for the Philosophy of Science, 55, 35-71.
Verzijden, M. N. & ten Cate, C. 2007. Early learning influences species assortative mating preferences in Lake Victoria cichlid fish. Biology Letters, 3, 134-136.
Pour en savoir plus (en français) :
Danchin, É., Giraldeau, L.-A., Valone, T. J. & Wagner, R. H. 2005. L’imitation dans le monde animal. Information publique et évolution culturelle. Terrain, 44, 91-108.
Jayat D., 2010. Les animaux ont-ils une culture ? EDP Sciences, 220p.
——
Article édité par Anne Teyssèdre
——
Absolument !
Je crois qu’il est important d’ajouter les symbiontes à la liste de ces supports à l’information génétique qui sont héritables et permettent l’adaptation de leur hôte (cf Fellous, Duron & Rousset, Nature Reviews Genetics, 2011). Rien que chez les arthropodes, une dizaine d’études ont récemment montré que l’acquisition et la transmission de symbiontes bactériens avait permis une évolution rapide du phénotype de l’hôte. Mais l’adaptation grâce aux symbiontes concerne probablement la majorité des eucaryotes. Les exemples les plus évidents sont naturellement les mitochondries et chloroplastes, qui dérivent d’anciennes bactéries, et dont les rôles pour la respiration et la photosynthèse ne sont plus à décrire.
Puisque nous sommes à souligner les différences entre information génétique (nucléaire) et symbiontes, soulignons 2 différences majeures: les symbiontes sont fréquemment acquis horizontalement depuis l’environnement tandis que les mutations des gènes nucléaires sont généralement le fruit du hasard ; de plus les symbiontes sont transmis par les mères beaucoup plus souvent que par les pères ce qui n’est pas le cas des gènes nucléaires.
Oui, je suis d’accord, et rien de ce qui est écrit ici n’exclut cette situation. L’acquisition de symbiontes transmis verticalement (c’est-à-dire de parent à enfant) est inclue dans l’hérédité génétique dont le véhicule est la séquence de la molécule de l’ADN du symbionte. Cela est vrai une fois que le symbionte est acquis. Mais aussi il me semble, pendant la phase d’acquisition où le futur symbionte n’est pas encore un symbionte, on peut imaginer que le gros de l’hérédité de l’ensemble Hôte-future symbionte est déjà véhiculé par l’ADN du symbionte et de l’hôte. Donc cela est clairement inclu dans l’hérédité élargie telle que nous la définissons ici. Qu’en penses tu? Par contre, c’est en effet un exemple intéressant et il est vrai que l’hérédité génétique ne se fait pas seulement verticalement, mais, contrairement à l’idée reçue, ceci est aussi le cas des gènes nucléaire, quoiqu’à des taux très différents..
Bonjour,
Merci pour cet article intéressant, dont la plus grande qualité est de dire tout haut, ce que beaucoup disent tout bas. J’aimerais ici attirer l’attention sur 3 points, qui ne viennent que renforcer les arguments de l’article.
1. Il eut été intéressant, dans les pages de la SFE, de retenir et d’illustrer l’hérédité « écologique » (notamment pour ceux qui, comme moi, n’auront pas le temps d’aller lire les articles originaux !-)
2. Bien que le choix des références d’un article soit éminemment subjectif, il me semble que le livre de Jablonka et Lamb (Evolution in four dimensions) est exactement dans le thème de l’article. Bien que ce livre soit parfois discutable, il présente une synthèse intéressante de la question.
3. Il devient urgent de discuter, dans la communauté des écologues comme dans ces pages de la SFE, la question de la possible « évolution » des écosystèmes. En effet, avec l’idée qu’un écosystème est fait de matériel génétique comme non génétique, on ne doute plus qu’il présente une certaine forme d’hérédité également. Ce sont sans doute les notions de parents et descendants qui prêtent plus à confusion, et qu’il faudrait faire l’effort de redéfinir. Par contre, avec la notion de « variant », je ne doute pas que l’on se rapproche insensiblement de l’idée que les écosystèmes « évoluent ». Avec cette propriété, ils ne deviendraient de fait qu’une représentation extrême (comme le rappelle la première phrase de l’article) de la Vie.
Oui merci pour ces trois commentaires qui me paraissent très pertinents.
1- J’ai pour aller vite et pour des raisons de place préféré n’illustrer que les deux aspects qui sont aux extrémités d’un gradient depuis ce que certains appellent « hard to soft inheritance ». Le mieux est de lire l’article dont s’inspire ce regard, toutes les références y sont.
2- Le livre de Jablonka et Lamb n’est pas cité dans le regard, mais aurait pu/du y être. Il est cité en position 130 dans l’article original avec d’autres articles importants de cet auteur.
3- Je ne suis pas certain de tout comprendre dans l’idée, mais il me semble que c’est à creuser en effet. On aborde des questions dans ce sens dans l’article original lorsque l’on remarque que ces processus non génétiques font émerger d’autres niveaux de sélection. Nous parlons dans ce cadre rapidement du fait que cela explique très bien la sélection de groupe par exemple. Ton idée pousse encore plus loin et cela me semble pertinent, même si elle reste encore à préciser. Il ne faut pas se limiter aux cadres un peu trop dogmatiques comme c’est malheureusement trop le cas avec la vision génocentrée actuelle. Donc, si tu veux défendre une réflexion dans ce sens, je suis très curieux de connaître où cela peut conduire. Ce serait intéressant à soumettre ces idées à des gens travaillant sur les dynamique écologiques et évolutives des communautés. Peut-être a t’elle déjà émergé, je ne sais pas à vrai dire.
A propos des symbioses et d’hérédité au sens large : Il me semble que la naissance et l’évolution de symbioses entre deux ou plusieurs espèces, auparavant indépendantes l’une de l’autre, illustrent très bien l’importance de l’interaction entre les hérédités génétique et écologique.
L’évolution d’une association symbiotique entre deux (ou plusieurs) organismes peut en effet être décrite comme la modification adaptative (génétique et écologique) de chacun des partenaires, en réponse aux changements des conditions et besoins écologiques liés à la présence du ou des autres partenaires, au fil des générations. Il s’agit en somme d’une « co-construction » de niches, au terme de laquelle les niches écologiques de deux partenaires peuvent éventuellement fusionner pour former la niche (écologique) d’une seule et même espèce de type « chimère », qui peut (si sa valeur adaptative est élevée) s’avérer l’espèce fondatrice d’une nouvelle lignée composite – algue primaire, secondaire ou tertiaire, animal capable de chimiosynthèse tel que les pogonophores/riftias, etc. – hébergeant dans ces cellules des microorganismes (bactéries, algues primaires..) voire des organites (mitochondires, chloroplastes) descendants de microoganismes symbiotiques…
Chez les espèces encore distinctes formant des associations symbiotiques, la séparation expérimentale ou « naturelle » des partenaires peut être vue comme un bouleversement de l’hérédité écologique, auquel le partenaire isolé, incomplet au plan physiologique (génétique et écologique), ne survit pas : c’est le cas pour le blanchiment et la disparition massive actuelle des coraux, par perte de leurs algues symbiotiques (zooxanthelles), en réponse au réchauffement et à l’acidification de l’eau des océans en région tropicale. De même, bien des champignons disparaissent faute de leur partenaire écologique héréditaire (arbre d’une certaine espèce), et les termites privés de leurs partenaires héréditaires protozoaires s’étiolent faute de digérer la cellulose…
Les symbioses impliquant un partenaire animal, enfin, peuvent relever non seulement de l’hérédité génétique et écologique, mais aussi parentale voire culturelle pour les vertébrés : c’est en têtant leur mère que les veaux acquièrent les bactéries symbiotiques qui autoriseront plus tard leur digestion d’herbe, et c’est (peut-être) en imitant le comportement coprophage des adultes que les jeunes lapins complètent leur flore intestinale de bactéries et protistes, digérant « pour eux » la cellulose.
Étienne, Anne,
Il me semble qu’à vous deux vous mettez justement le doigt sur la particularité de l’évolution grâce aux symbiontes: en partie elle relève du bon vieil ADN, mais aussi de l’hérédité écologique et même culturel.
De façon plus générale, peut être que ce qui se profile à l’horizon c’est un re-découpage des caractéristiques qui identifient tel ou tel type d’évolution. Autrement dit, si évolutions génétiques, écologiques, culturelles, etc… ont tellement en commun c’est peut être que nous n’avons pas encore identifié la logique et les facteurs qui nous permettrons de mettre de l’ordre et du sens dans cette diversité de processus.
Simon, Je ne vois pas comment l’hérédité culturelle peut être impliquée dans une relation symbiotique telle que celle des lichens par exemple. On donne une définition bien précise de l’hérédité culturelle qui implique une certaine forme de cognition et d’interaction sociale qui peut difficilement s’appliquer à des microorganismes. Je suppose que tu fais allusion à des symbioses antre macroorganismes.
De même, dans les commentaires d’Anne ci-après, j’ai l’impression que l’on ne parle pas de la même chose par moment au sujet de l’hérédité écologique. C’est dur de faire cela par messages courts de la sorte.
Anne, pour ce qui est du côté révolutionnaire ou non, tout dépend aussi de comment on regarde les choses. Tu cites le livre de Olding-Smee et al. C’est typiquement le genre d’ouvrage qui montre justement à quel point la prise en compte de l’hérédité non génétique change toutes les règles du jeu au point que des situations impossibles dans le cadre de la synthèse moderne deviennent tout à fait explicables dans le cadre d’une vision plus généralisée. Je pense que dans les faits le saut conceptuel est très grand et que la théorie généralisée qui en ressortira sera aussi novatrice que l’a été la relativité par rapport à la vision Newtonienne.
Dans ce vaste mouvement de fond qui a commencé à la fin des années 70, ce que nous avons fait se réduit à une chose, mais elle est de taille: nous reformulons tous les arguments apportés par de nombreux auteurs en termes d’hérédité et d’héritabilité. Du coup, cela ouvre la possibilité d’utiliser tous les outils de la génétique quantitative pour étudier la variation héritable non génétique. C’est tout ce que nous faisons, mais je pense que c’est une avancée importante.
Bonjour,
Personnellement, je ne crois pas qu’il s’agisse là d’une révolution de la théorie de l’évolution, mais de développements et prolongements de cette théorie, avec l’exploration des interactions entre facteurs génétiques, épigénétiques, sociaux et culturels impliqués dans l’évolution des organismes.
– « L’hérédité écologique » (bien que non désignée dans ces termes) est implicite dans et à la base même de la théorie darwinienne de l’évolution des espèces au moyen de la sélection naturelle (et de la sélection sexuelle) : l’adaptation des populations à leurs conditions de vie, ou des différentes stratégies sexuelles mâles et femelles à l’intérieur des populations, présupposent en effet une certaine stabilité/« héritabilité » de ces conditions de vie et de reproduction au fil des générations – conditions de vie qui sont en retour modifiés au fil du temps par l’évolution des espèces, en interaction entre elles et avec leur milieu de vie.
(- Dans leur livre « Niche construction » daté de 2003, Olding-Smee, Laland et Feldman ont enrichi la théorie de l’évolution en soulignant l’importance potentielle de l’impact-retour des espèces sur leurs conditions de vie, et en explorant la dynamique évolutive de l’interaction entre les uns et les autres, au fil des générations.)
– L’interaction entre génome et culture, sélection naturelle et culturelle, est reconnue et explorée avec succès depuis au moins une trentaine d’années (Cavalli Sforza et Feldman, 1981) ;
– Le rôle de l’épigénèse dans l’évolution des conflits parents-progéniture a été souligné par David Haig au milieu des années 1980 mais relativement peu étudié depuis…
– L’interaction entre le génome, les facteurs épigénétiques et l’environnement, enfin, renvoie à la notion de « phénotype étendu » proposé par Richard Dawkins en 1982.
Il me semble par ailleurs important (et même nécessaire, face aux changements globaux actuels…), comme à Cédric G., de compléter/situer la théorie « multifactorielle » de l’évolution des espèces par l’exploration des mécanismes d’évolution des communautés et écosystèmes…
Bonjour et meilleurs voeux à tous,
Merci beaucoup pour cet intéressant exposé. Je voulais juste mentionner que l’évolution des écosystèmes (ou plutôt des communautés, assemblages ou populations) est un concept qui a déjà été présenté plusieurs fois. L’exemple que je connais est le papier de Swenson et al. 2000 dans PNAS. L’idée est néanmoins vivement critiquée par certain. Personnellement je la trouve plutôt stimulante, et j’imagine que les apprentissages de part et d’autres des champs écologie des communautés / écologie évolutive pourrait gagner à être confrontés (comme Venner et al, 2009 Trends in genetics).
Donc cette idée a déjà une histoire. Très bien. Je trouve que l’on a tendance à vouloir ignorer les unités de sélection supérieures à l’individu; certains d’ailleurs ne comprennent pas comment on peut parler d’unité de sélection au-dessus du gène. Mais bon, ils sont plutôt rares aujourd’hui il me semble. Pourtant historiquement, la plupart des transitions majeures de l’évolution ont du impliquer des formes de transmission non purement génétique. Ce fut très probablement le cas pour celle concernant l’émergence des organismes métacellulaires.
Donc tout un champ à explorer en effet.
Bonjour,
Il y a aussi des articles intéressants et plus vieux d’écologie évolutive sur ce sujet, de Geraat Vermeij, publiés dans les années 1980. (Et voir son livre Evolution and Escalation: An Ecological History of Life, publié par Princeton University Press en 1987.)
Et dans un autre registre, avec une approche systémique appliquée aux changements globaux actuels, il y a aussi la théorie des cycles (« Panarchie ») de L.H. Gunderson et C.S. Holling (cf leur livre Panarchy, 2004, Island Press)…
[Mais ce regard d’Etienne D. est centré sur l’hérédité non génétique dans les populations et espèces, un sujet déjà passionnant et complexe…]
Anne
Bonjour,
Merci pour cet article. Le mot culturel étant déjà porteur de définitions en Sciences Humaines, je pense qu’il faudrait compléter les types d’hérédité présentés ici pour éviter un effet fourre-tout. En plus de l’hérédité culturelle qui serait réservée à l’homme et à certaines espèces animales, il serait alors pertinent de définir aussi une hérédité sociale ou une hérédité comportementale ou éthologique. Amicalement.